

Review
doi: 10.1016/j.cell.2013.11.011.
Incisive imaging and computation for cellular mysteries: lessons from abscission
Affiliations
- PMID: 24315094
- PMCID: PMC4220545
- DOI: 10.1016/j.cell.2013.11.011
Item in Clipboard
Review
Incisive imaging and computation for cellular mysteries: lessons from abscission
Cell.
.
Abstract
The final cleavage event that terminates cell division, abscission of the small, dense intercellular bridge, has been particularly challenging to resolve. Here, we describe imaging innovations that helped answer long-standing questions about the mechanism of abscission. We further explain how computational modeling of high-resolution data was employed to test hypotheses and generate additional insights. We present the model that emerges from application of these complimentary approaches. Similar experimental strategies will undoubtedly reveal exciting details about other underresolved cellular structures.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
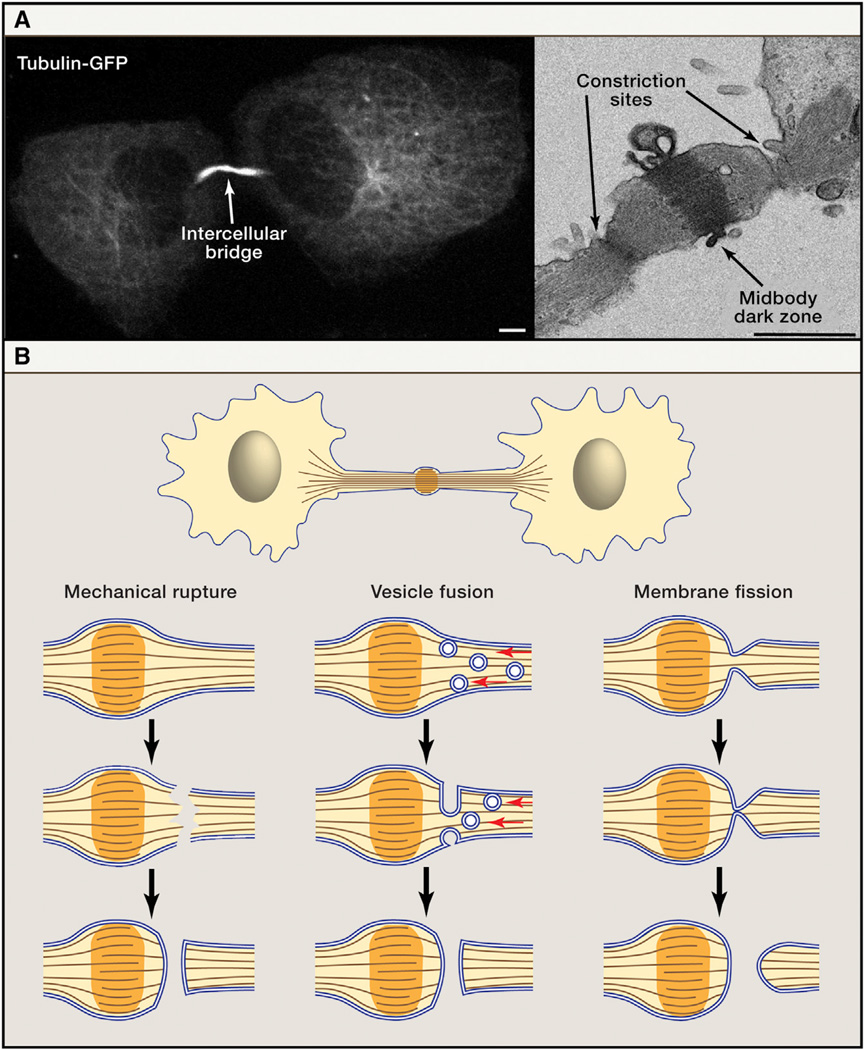
(A) The intercellular bridge connecting two daughter cells must be resolved to complete cytokinesis. The microtubules in the intercellular bridge are visible using both confocal microscopy and TEM. Shown here are live MDCK cells expressing tubulin-GFP (left) and fixed MDCK cells imaged by TEM (right) (Elia et al., 2011). The bridge can be severed on either or both sides of the dark zone. Scale bars: (left) 5 µm, (right) 1 µm. (B) Schematic representation of the three classic models for abscission of the intercellular bridge. The mechanical rupture model (left) posited that as daughter cells move away from one another, the intercellular bridge is torn. The opening is then sealed by cellular wound healing mechanisms. The vesicle fusion model (center) proposed that vesicles delivered to the midbody fuse with each other and with the plasma membrane to separate the daughter cells. Themembrane fission model (right) suggested that acute reduction in the diameter of the bridge membrane brings the lipid bilayers close enough to fuse. Prior to the technical innovations discussed in this review, poor spatial and temporal resolution made it difficult to differentiate between these models experimentally.
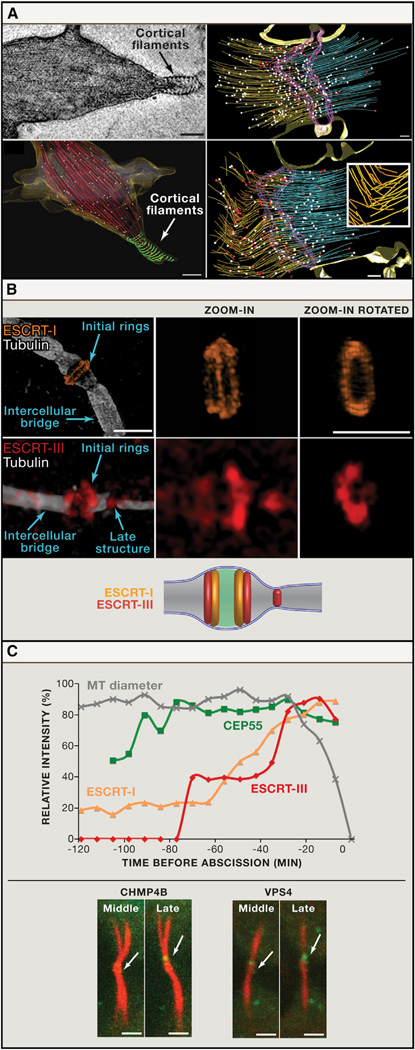
(A) Cryo-EM tomography of the intercellular bridge. Left: the identification of cortical filaments at the membrane of the intercellular bridge in cells at late cytokinesis. The original cryo-EM image (top) and a 3D reconstruction of the entire tomogram series (bottom) are shown. Red: microtubules, green: cortical filaments. Scale bar, 200 nm. Images are from Guizetti et al. (2011) with permission. Right: Reconstructions of EM tomogram from cells at early (top) and late telophase (bottom). The number of microtubules with flared ends indicative of rapidly depolymerizing microtubules (red dots), increases in late telophase (relative to microtubules with nonflared ends; white dots). Also note the appearance of buckled microtubules in cells at late telophase. (Purple: midbody dark zone; yellow and blue: microtubules). Scale bar, 5 µm. Inset: enlarged view from the buckled microtubules zone. Images are Schiel et al. (2011) with permission. (B) Structural organization of ESCRT complex components at the intercellular bridge as revealed by SIM. ESCRT-I component TSG101 (top) localizes to ring structures on either side of the dark zone. The ESCRT-III protein CHMP4B (bottom) is visible both in rings adjacent to the dark zone and at a late structure closer to the cell body. An integrated model of ESCRT localization based on measurements from SIM images is shown below. Scale bar, 2 µm. (C) Kinetics of cytokinetic abscission assessed using high-speed quantitative fluorescence imaging. ESCRT components are recruited to the intercellular bridge at different times prior to abscission (time 0). As shown in the graph, the relative intensity values CEP55 increases early. ESCRT-I and ESCRT-III levels increase later in abscission. An acute increase in ESCRT-III levels correlates with a decrease in the diameter of the intercellular bridge. Lower panel: representative middle and late-phase images from time series of cells expressing either the ESCRT-III protein CHMP4B or the AAA-ATPase VPS4. Note that both CHMP4B and VPS4 have similar localization patterns (arrows). Scale bar, 2 µm.
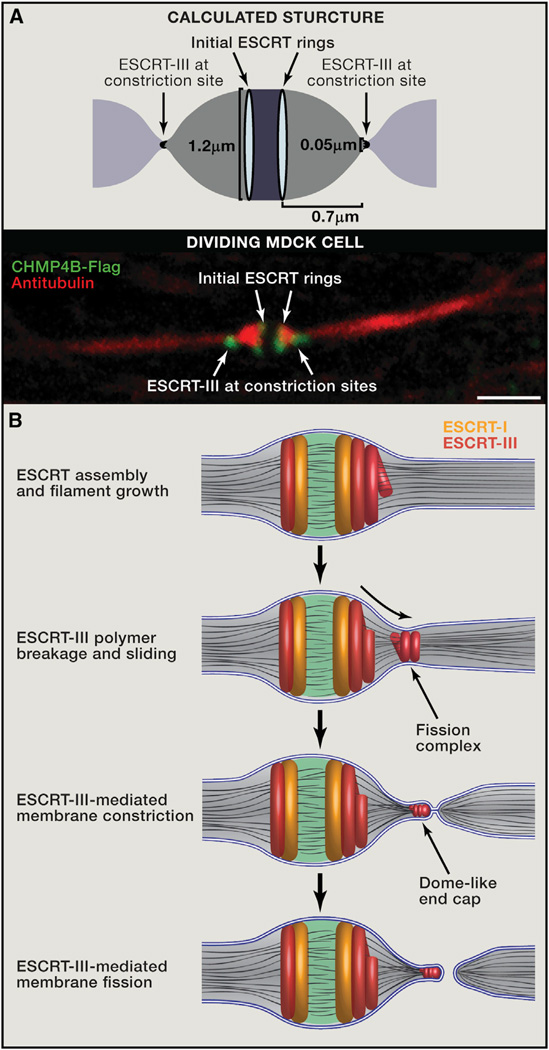
(A) The calculated shape of the intercellular bridge from computational modeling (top) and the morphology of the bridge as observed by tubulin and ESCRT-III staining using spinning disk confocal microscopy in dividing MDCK cells (bottom) are very similar. The model predicts that when the bridge diameter is 1.2 µm, the optimal distance between the initial and late ESCRT-III pools will be 0.7 µm, which is similar to the distance measured experimentally. Scale bar, 2 µm. (B) Suggested mechanistic model for ESCRT-driven cytokinetic abscission based on high-resolution microscopy data and computational modeling. As detailed in the text, cytokinetic abscission begins with the assembly of early and late ESCRT proteins into a series of partially overlapping cortical rings located at the center of the intercellular bridge. Ring formation is followed by ESCRT-III polymerization and remodeling into 3D helical spirals. Breakage and sliding of the membrane-associated ESCRT-III spiral away from the dark zone, results in acute constriction of the cytokinetic tube. This continues until the ESCRT-III spiral reaches an equilibrium distance where it relaxes to a spontaneous diameter of 50 nm. At this point fission of the 50 nmdiameter constricted membrane tube occurs, mediated by a dome-like end-cap structure, finalizing cell separation. Similar events occur on the other side of the bridge (not illustrated).
References
-
- Barr FA, Gruneberg U. Cytokinesis: placing and making the final cut. Cell. 2007;131:847–860. - PubMed
-
- Carlton JG, Martin-Serrano J. Parallels between cytokinesis and retroviral budding: a role for the ESCRT machinery. Science. 2007;316:1908–1912. - PubMed
-
- Dambournet D, Machicoane M, Chesneau L, Sachse M, Rocancourt M, El Marjou A, Formstecher E, Salomon R, Goud B, Echard A. Rab35 GTPase and OCRL phosphatase remodel lipids and F-actin for successful cytokinesis. Nat. Cell Biol. 2011;13:981–988. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
