

Circadian control of global gene expression by the cyanobacterial master regulator RpaA
- PMID: 24315105
- PMCID: PMC3935230
- DOI: 10.1016/j.cell.2013.11.005
Circadian control of global gene expression by the cyanobacterial master regulator RpaA
Abstract
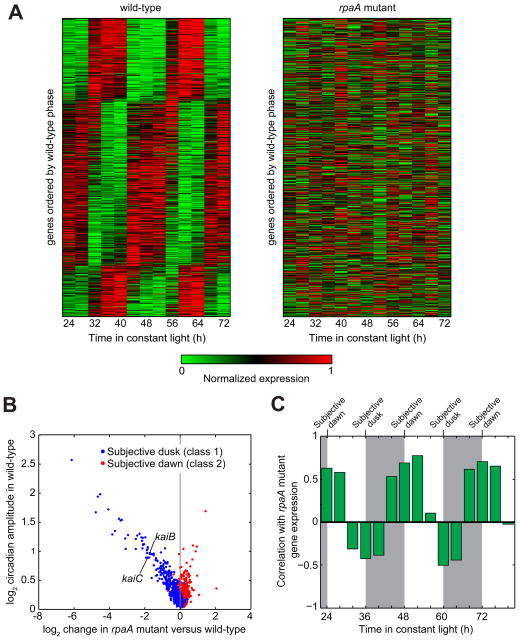
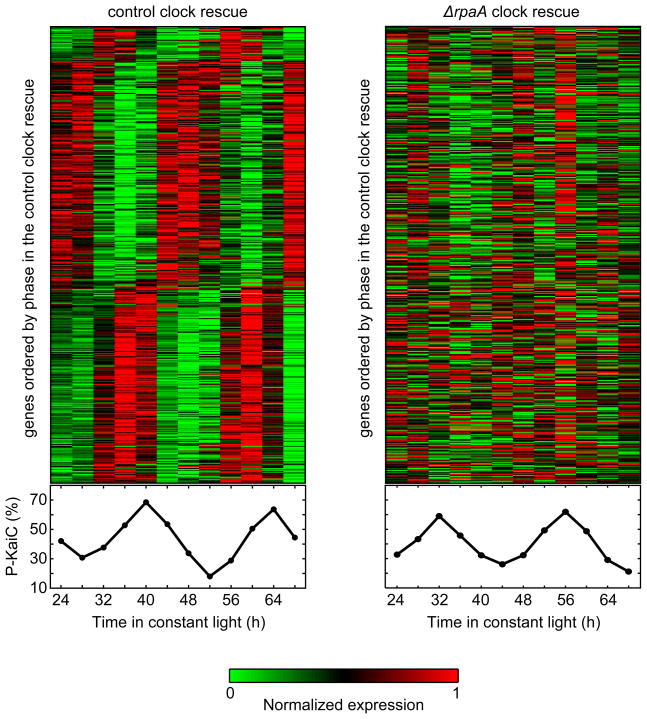
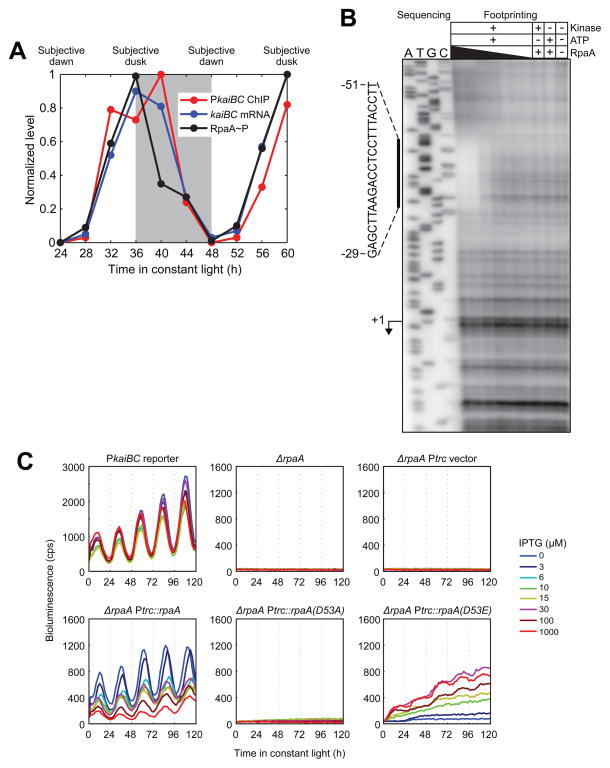
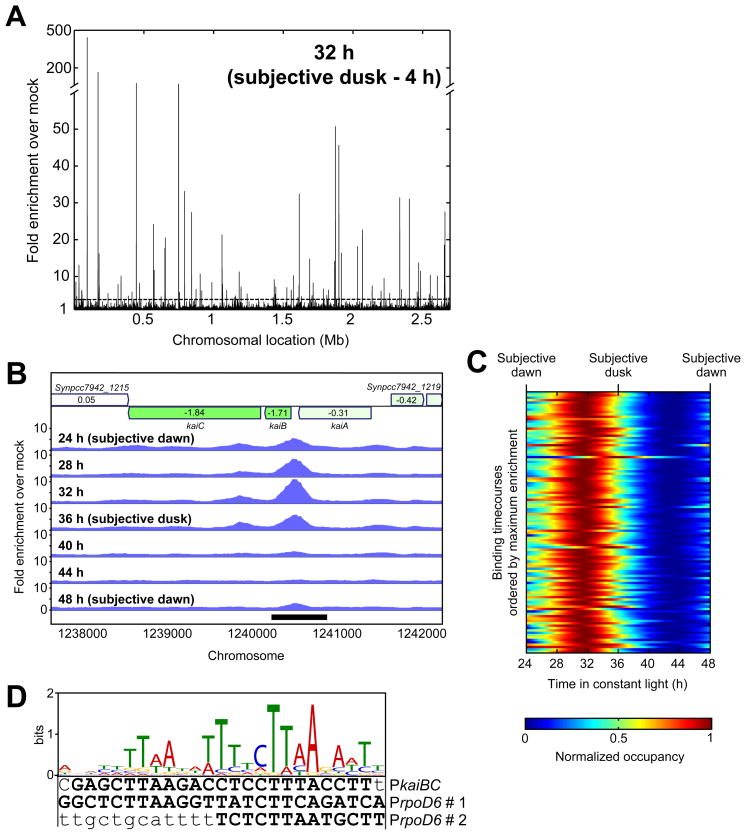
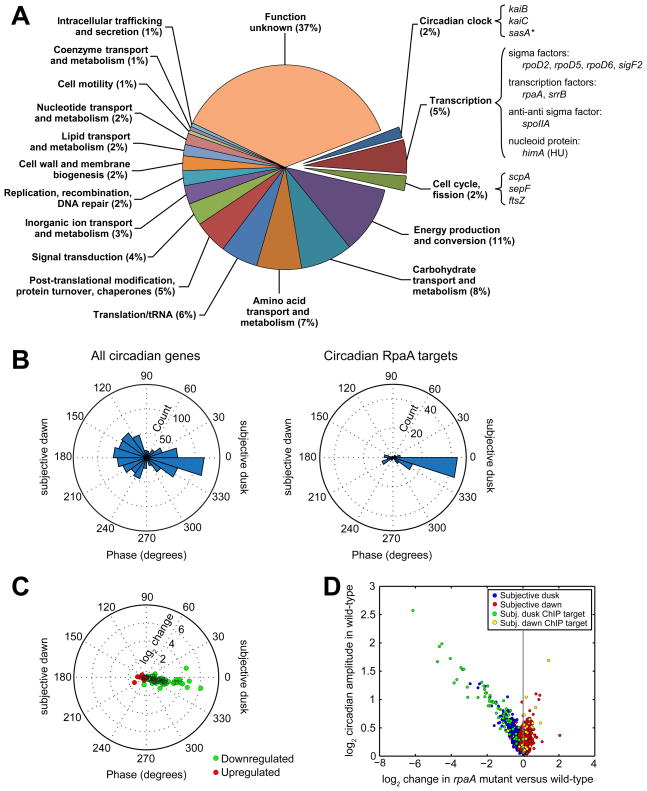
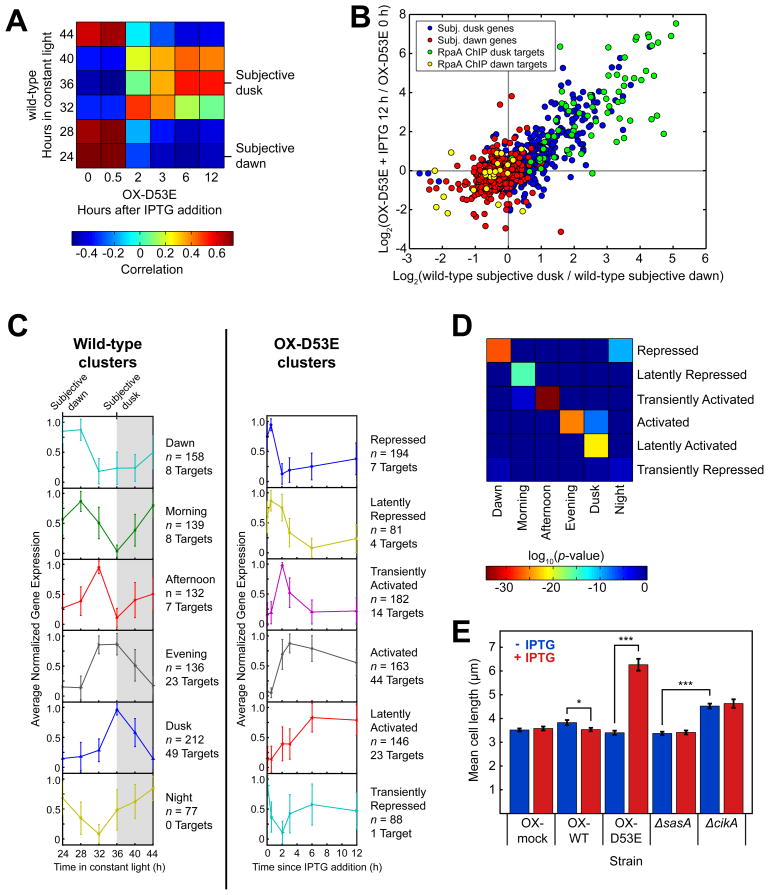
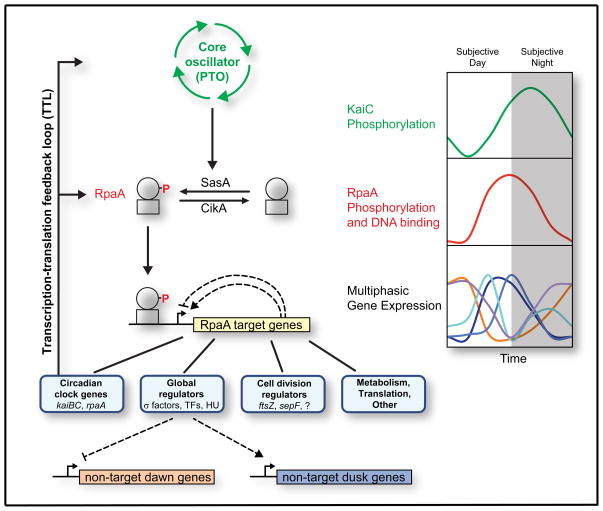
The cyanobacterial circadian clock generates genome-wide transcriptional oscillations and regulates cell division, but the underlying mechanisms are not well understood. Here, we show that the response regulator RpaA serves as the master regulator of these clock outputs. Deletion of rpaA abrogates gene expression rhythms globally and arrests cells in a dawn-like expression state. Although rpaA deletion causes core oscillator failure by perturbing clock gene expression, rescuing oscillator function does not restore global expression rhythms. We show that phosphorylated RpaA regulates the expression of not only clock components, generating feedback on the core oscillator, but also a small set of circadian effectors that, in turn, orchestrate genome-wide transcriptional rhythms. Expression of constitutively active RpaA is sufficient to switch cells from a dawn-like to a dusk-like expression state as well as to block cell division. Hence, complex global circadian phenotypes can be generated by controlling the phosphorylation of a single transcription factor.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Ashby MK, Mullineaux CW. Cyanobacterial ycf27 gene products regulate energy transfer from phycobilisomes to photosystems I and II. FEMS Microbiol Lett. 1999;181:253–260. - PubMed
-
- Ducat DC, Way JC, Silver PA. Engineering cyanobacteria to generate high-value products. Trends Biotechnol. 2011;29:95–103. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
