

Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module
- PMID: 24315106
- PMCID: PMC3925772
- DOI: 10.1016/j.cell.2013.11.004
Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module
Abstract
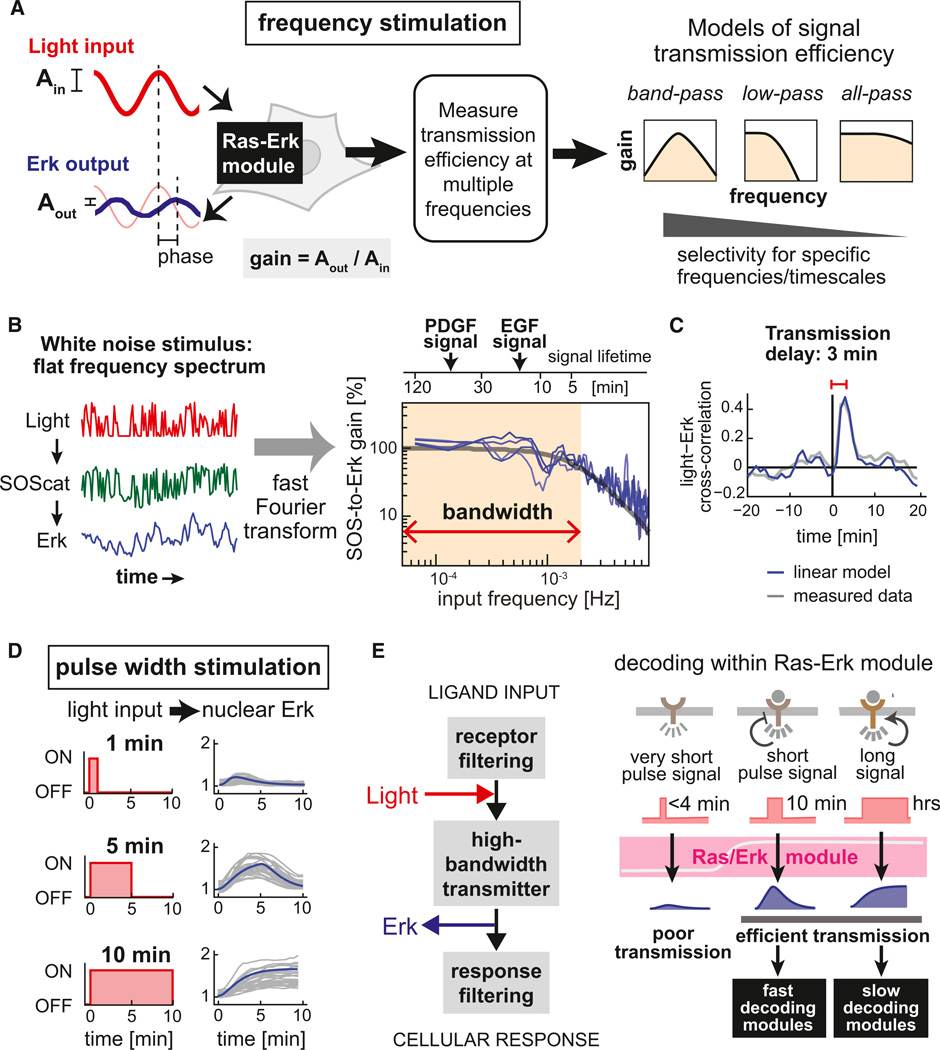
The complex, interconnected architecture of cell-signaling networks makes it challenging to disentangle how cells process extracellular information to make decisions. We have developed an optogenetic approach to selectively activate isolated intracellular signaling nodes with light and use this method to follow the flow of information from the signaling protein Ras. By measuring dose and frequency responses in single cells, we characterize the precision, timing, and efficiency with which signals are transmitted from Ras to Erk. Moreover, we elucidate how a single pathway can specify distinct physiological outcomes: by combining distinct temporal patterns of stimulation with proteomic profiling, we identify signaling programs that differentially respond to Ras dynamics, including a paracrine circuit that activates STAT3 only after persistent (>1 hr) Ras activation. Optogenetic stimulation provides a powerful tool for analyzing the intrinsic transmission properties of pathway modules and identifying how they dynamically encode distinct outcomes.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






References
-
- Amit I, Citri A, Shay T, Lu Y, Katz M, Zhang F, Tarcic G, Siwak D, Lahad J, Jacob-Hirsch J, et al. A module of negative feedback regulators defines growth factor signaling. Nat. Genet. 2007;39:503–512. - PubMed
-
- Barber MA, Donald S, Thelen S, Anderson KE, Thelen M, Welch HC. Membrane translocation of P-Rex1 is mediated by G protein betagamma subunits and phosphoinositide 3-kinase. J. Biol. Chem. 2007;282:29967–29976. - PubMed
-
- Bishop JM, Capobianco AJ, Doyle HJ, Finney RE, McMahon M, Robbins SM, Samuels ML, Vetter M. Proto-oncogenes and plasticity in cell signaling. Cold Spring Harb. Symp. Quant. Biol. 1994;59:165–171. - PubMed
-
- Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005;8:1263–1268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- GM55040/GM/NIGMS NIH HHS/United States
- P50 GM081879/GM/NIGMS NIH HHS/United States
- R01 GM062583/GM/NIGMS NIH HHS/United States
- R01 GM055040/GM/NIGMS NIH HHS/United States
- GM096164/GM/NIGMS NIH HHS/United States
- R01 GM096164/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- PN2 EY016546/EY/NEI NIH HHS/United States
- PN2EY016546/EY/NEI NIH HHS/United States
- R01 GM084040/GM/NIGMS NIH HHS/United States
- P50GM081879/GM/NIGMS NIH HHS/United States
- GM62583/GM/NIGMS NIH HHS/United States
- GM084040/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
