

Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation
- PMID: 24315368
- PMCID: PMC3939841
- DOI: 10.1016/j.cmet.2013.11.001
Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation
Abstract
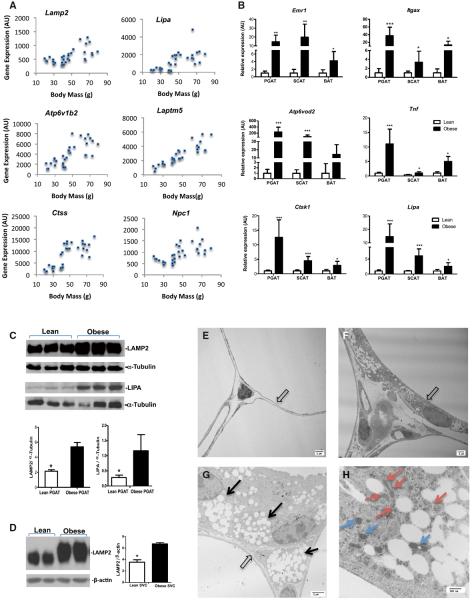
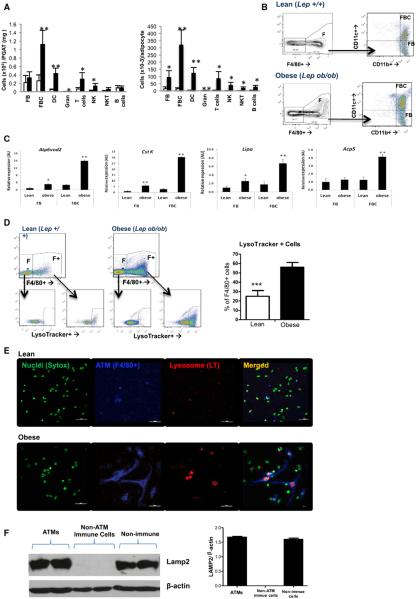
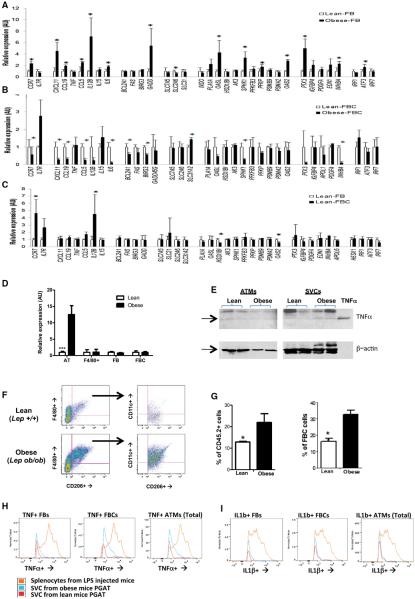
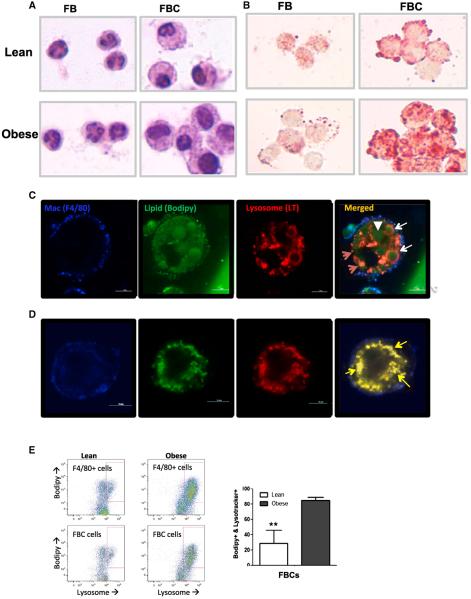
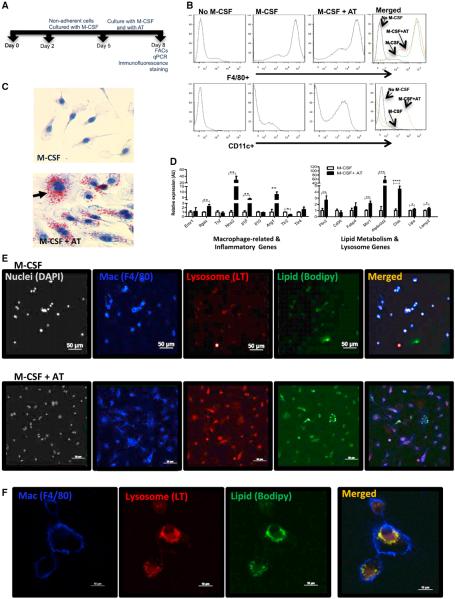
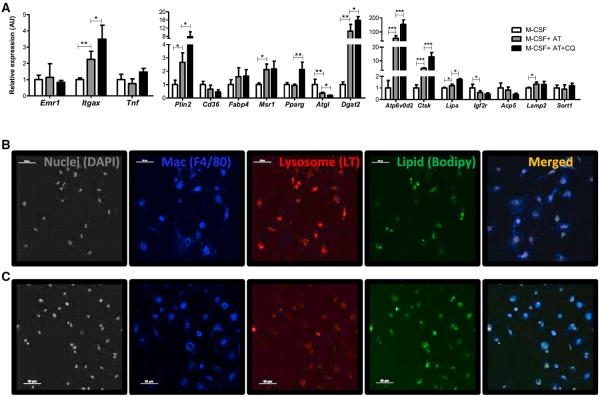
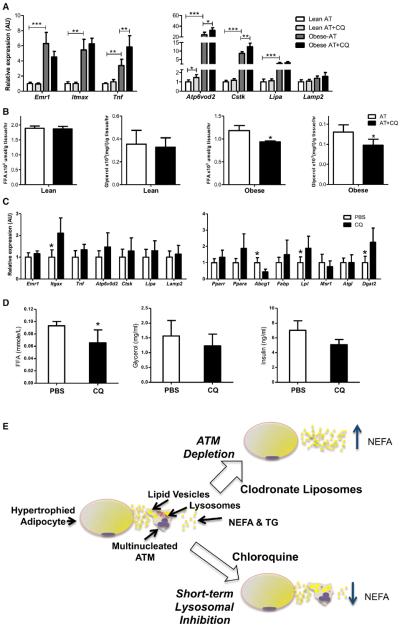
Obesity activates a complex systemic immune response that includes the recruitment of macrophages and other immune cells to key metabolic tissues. Current models postulate that obesity and excess lipids classically activate macrophages, polarizing them toward an M1 (inflammatory) state. Little is known about noninflammatory functions of adipose tissue macrophages (ATMs). Here, we show that the expansion of adipose tissue (AT) across models of obesity induces a program of lysosome biogenesis in ATMs and is associated with lipid catabolism but not a classic inflammatory phenotype. This program is induced by factors produced by AT and is tightly coupled to lipid accumulation by ATMs. Inhibition of ATM lysosome function impairs lipid metabolism and increases lipid content in ATMs and reduces whole AT lipolysis. These data argue that ATMs contribute quantitatively to the development of obesity-induced inflammation but also serve an important role in lipid trafficking independent of their inflammatory phenotype.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Adipose tissue macrophages: Amicus adipem?Cell Metab. 2013 Dec 3;18(6):767-8. doi: 10.1016/j.cmet.2013.11.011. Cell Metab. 2013. PMID: 24315364 Free PMC article.
-
Obesity: A complex role for adipose tissue macrophages.Nat Rev Endocrinol. 2014 Apr;10(4):193-4. doi: 10.1038/nrendo.2014.12. Epub 2014 Feb 4. Nat Rev Endocrinol. 2014. PMID: 24492182 No abstract available.
References
-
- Benoit FL. The inhibitory effect of chloroquine on rat adipose tissue metabolism in vitro. Metabolism. 1967;16:557–561. - PubMed
-
- Bourlier V, Zakaroff-Girard A, Miranville A, De Barros S, Maumus M, Sengenes C, Galitzky J, Lafontan M, Karpe F, Frayn KN, Bouloumié A. Remodeling phenotype of human subcutaneous adipose tissue macrophages. Circulation. 2008;117:806–815. - PubMed
-
- Edwards JR, Mundy GR. Advances in osteoclast biology: old findings and new insights from mouse models. Nat Rev Rheumatol. 2011;7:235–243. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
