

Circulating angiopoietin-like 4 links proteinuria with hypertriglyceridemia in nephrotic syndrome
- PMID: 24317117
- PMCID: PMC4114723
- DOI: 10.1038/nm.3396
Circulating angiopoietin-like 4 links proteinuria with hypertriglyceridemia in nephrotic syndrome
Abstract
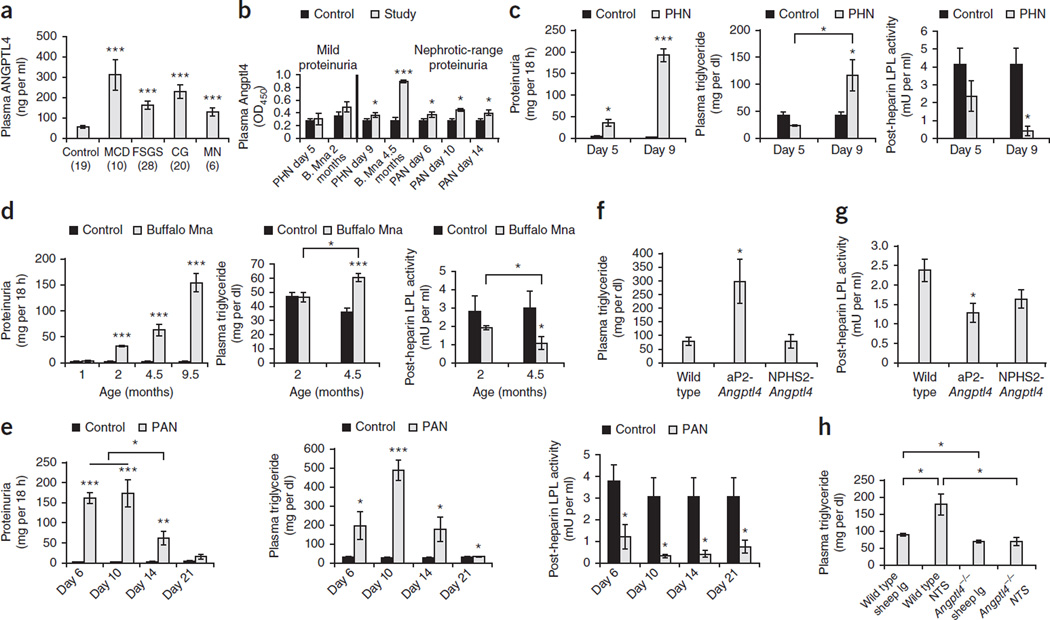
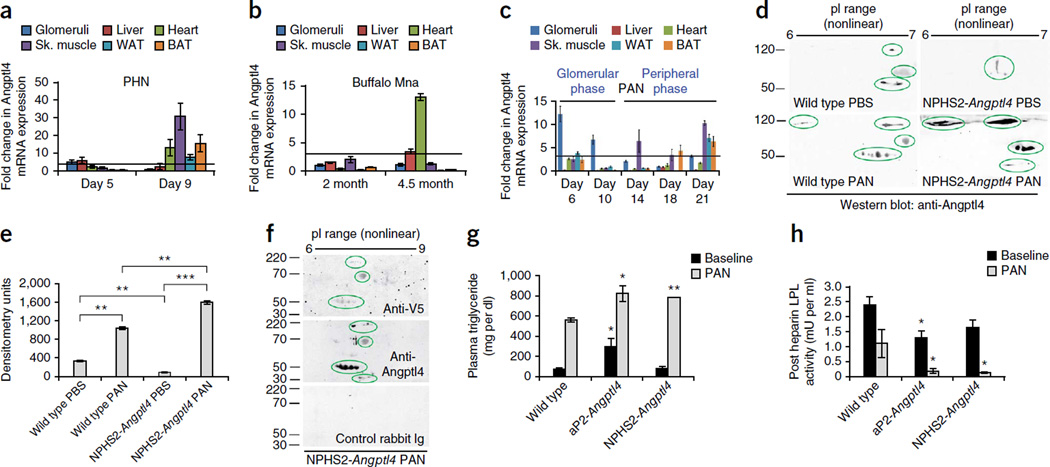
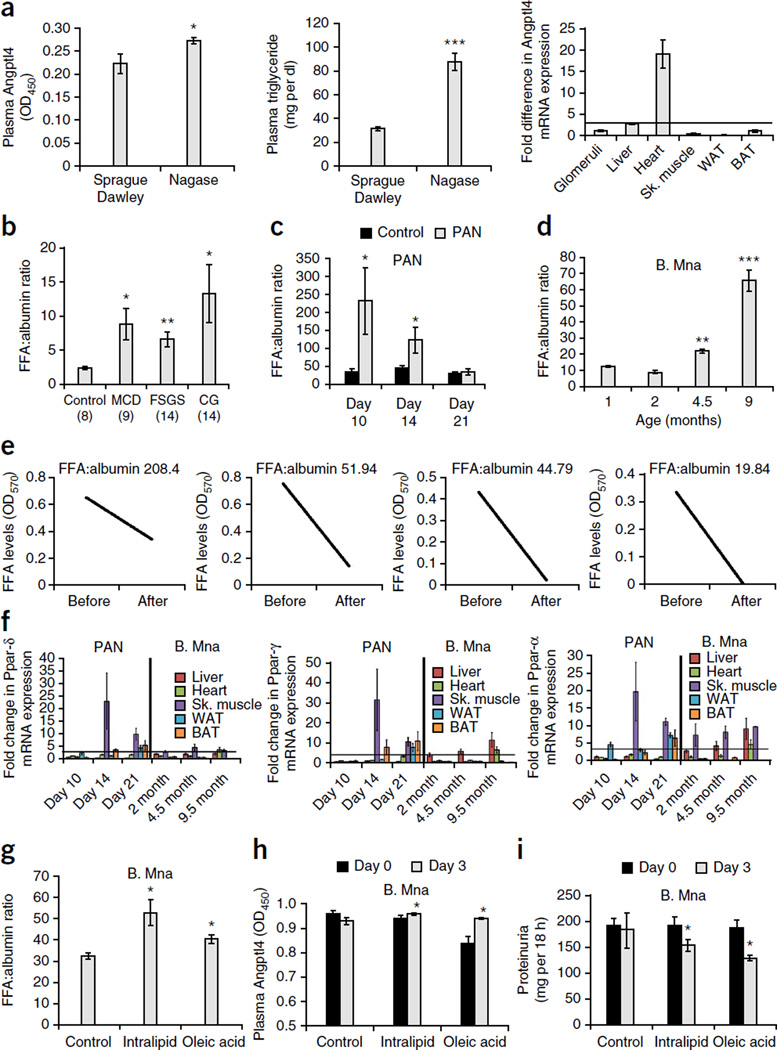
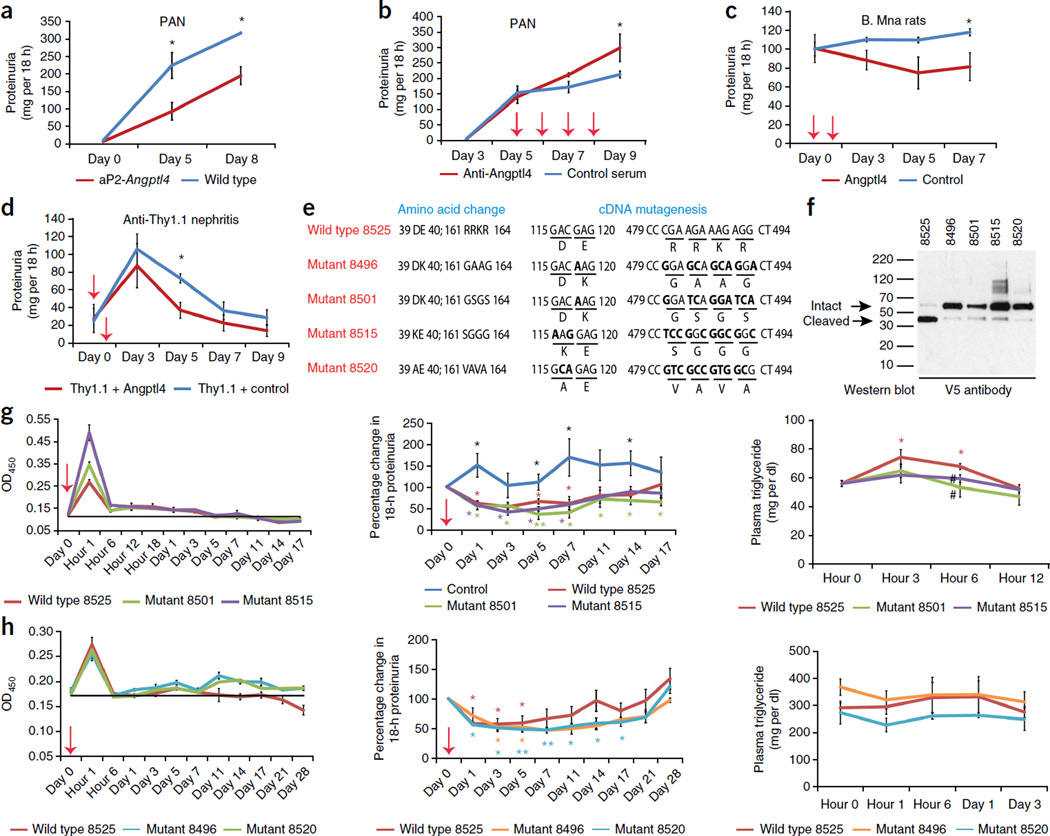
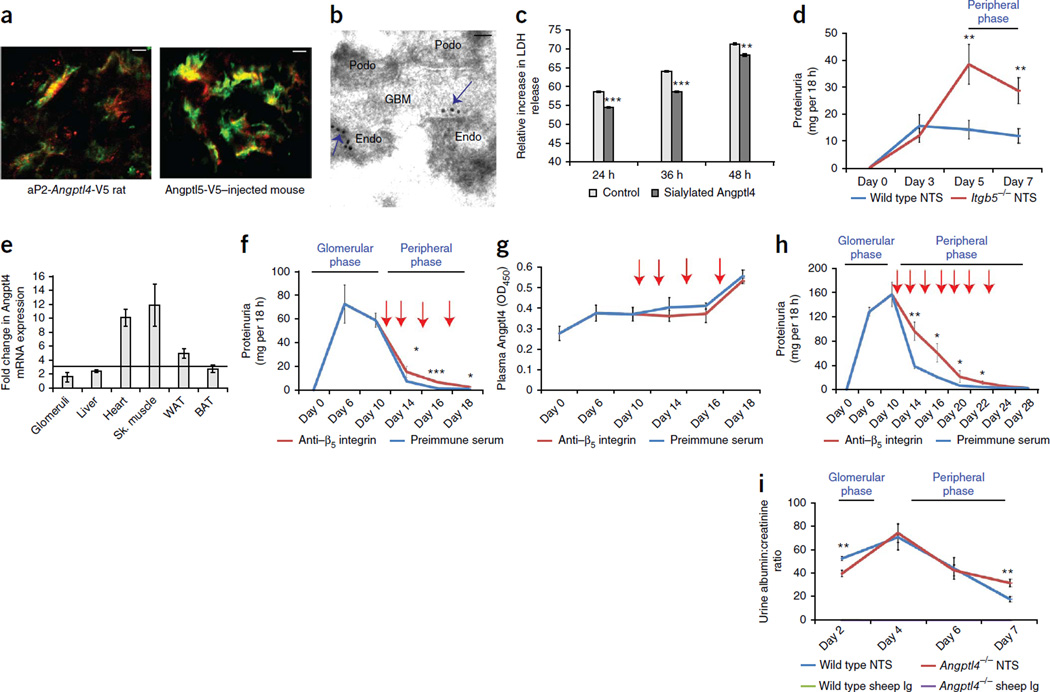
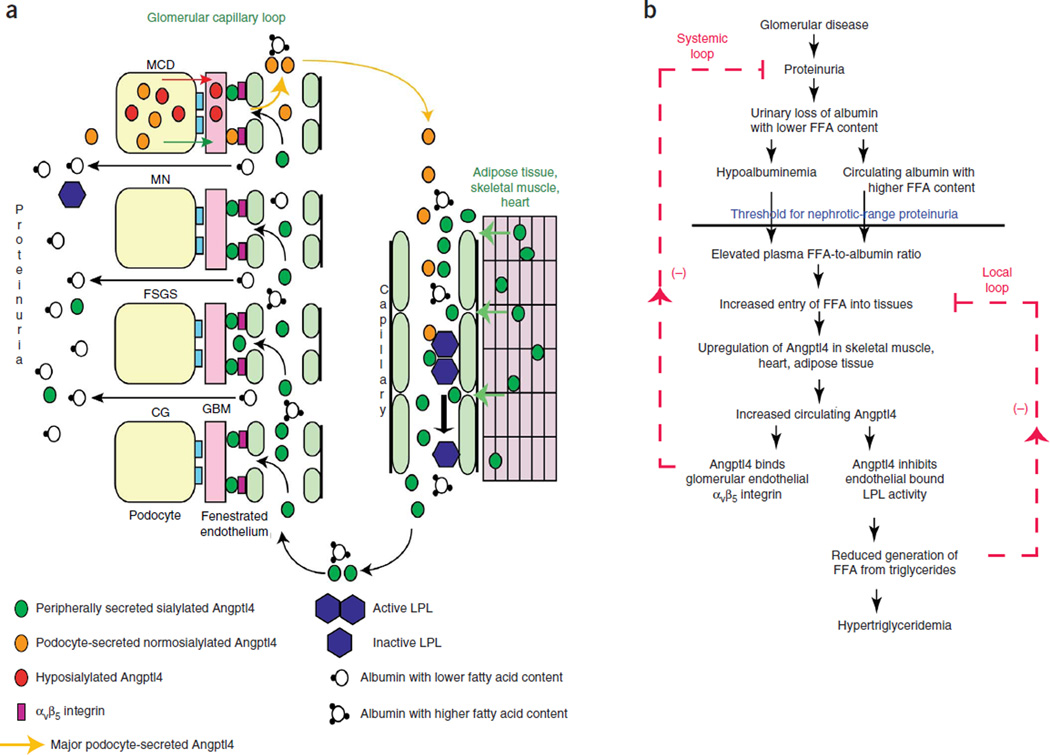
The molecular link between proteinuria and hyperlipidemia in nephrotic syndrome is not known. We show in the present study that plasma angiopoietin-like 4 (Angptl4) links proteinuria with hypertriglyceridemia through two negative feedback loops. In previous studies in a rat model that mimics human minimal change disease, we observed localized secretion by podocytes of hyposialylated Angptl4, a pro-proteinuric form of the protein. But in this study we noted high serum levels of Angptl4 (presumably normosialylated based on a neutral isoelectric point) in other glomerular diseases as well. Circulating Angptl4 was secreted by extrarenal organs in response to an elevated plasma ratio of free fatty acids (FFAs) to albumin when proteinuria reached nephrotic range. In a systemic feedback loop, these circulating pools of Angptl4 reduced proteinuria by interacting with glomerular endothelial αvβ5 integrin. Blocking the Angptl4-β5 integrin interaction or global knockout of Angptl4 or β5 integrin delayed recovery from peak proteinuria in animal models. But at the same time, in a local feedback loop, the elevated extrarenal pools of Angptl4 reduced tissue FFA uptake in skeletal muscle, heart and adipose tissue, subsequently resulting in hypertriglyceridemia, by inhibiting lipoprotein lipase (LPL)-mediated hydrolysis of plasma triglycerides to FFAs. Injecting recombinant human ANGPTL4 modified at a key LPL interacting site into nephrotic Buffalo Mna and Zucker Diabetic Fatty rats reduced proteinuria through the systemic loop but, by bypassing the local loop, without increasing plasma triglyceride levels. These data show that increases in circulating Angptl4 in response to nephrotic-range proteinuria reduces the degree of this pathology, but at the cost of inducing hypertriglyceridemia, while also suggesting a possible therapy to treat these linked pathologies.
Figures
Comment in
-
Nephrotic syndrome: Negative feedback loop reveals novel potential therapy.Nat Rev Nephrol. 2014 Feb;10(2):63. doi: 10.1038/nrneph.2013.267. Epub 2013 Dec 17. Nat Rev Nephrol. 2014. PMID: 24342959 No abstract available.
-
Dual role of circulating angiopoietin-like 4 (ANGPTL4) in promoting hypertriglyceridemia and lowering proteinuria in nephrotic syndrome.Am J Kidney Dis. 2014 Oct;64(4):495-8. doi: 10.1053/j.ajkd.2014.04.016. Epub 2014 May 16. Am J Kidney Dis. 2014. PMID: 24838183 No abstract available.
References
-
- Vaziri ND. Molecular mechanisms of lipid disorders in nephrotic syndrome. Kidney Int. 2003;63:1964–1976. - PubMed
-
- Marsh JB, Drabkin DL. Experimental reconstruction of metabolic pattern of lipid nephrosis: key role of hepatic protein synthesis in hyperlipemia. Metabolism. 1960;9:946–955. - PubMed
-
- Merkel M, Eckel RH, Goldberg IJ. Lipoprotein lipase: genetics, lipid uptake, and regulation. J. Lipid Res. 2002;43:1997–2006. - PubMed
-
- Shearer GC, Kaysen GA. Endothelial bound lipoprotein lipase (LpL) depletion in hypoalbuminemia results from decreased endothelial binding, not decreased secretion. Kidney Int. 2006;70:647–653. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
