

Identification of residues required for stalled-ribosome rescue in the codon-independent release factor YaeJ
- PMID: 24322300
- PMCID: PMC3950681
- DOI: 10.1093/nar/gkt1280
Identification of residues required for stalled-ribosome rescue in the codon-independent release factor YaeJ
Abstract
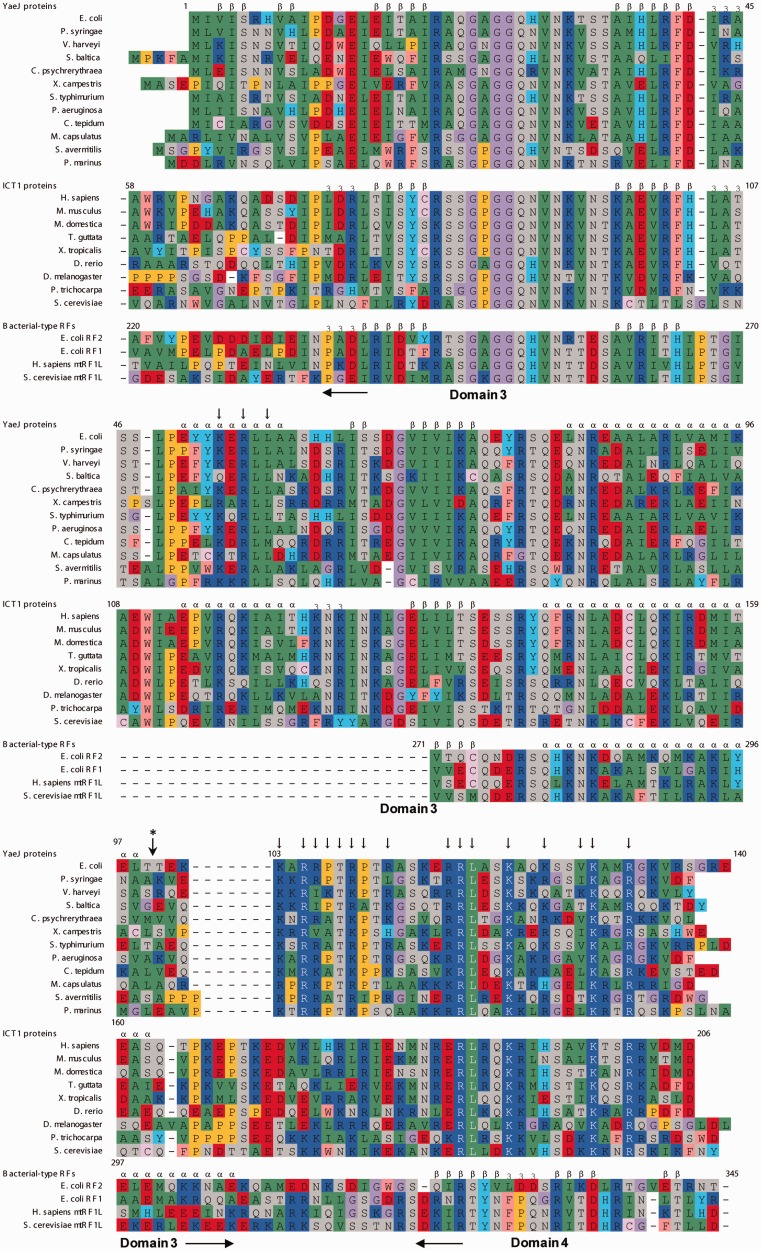
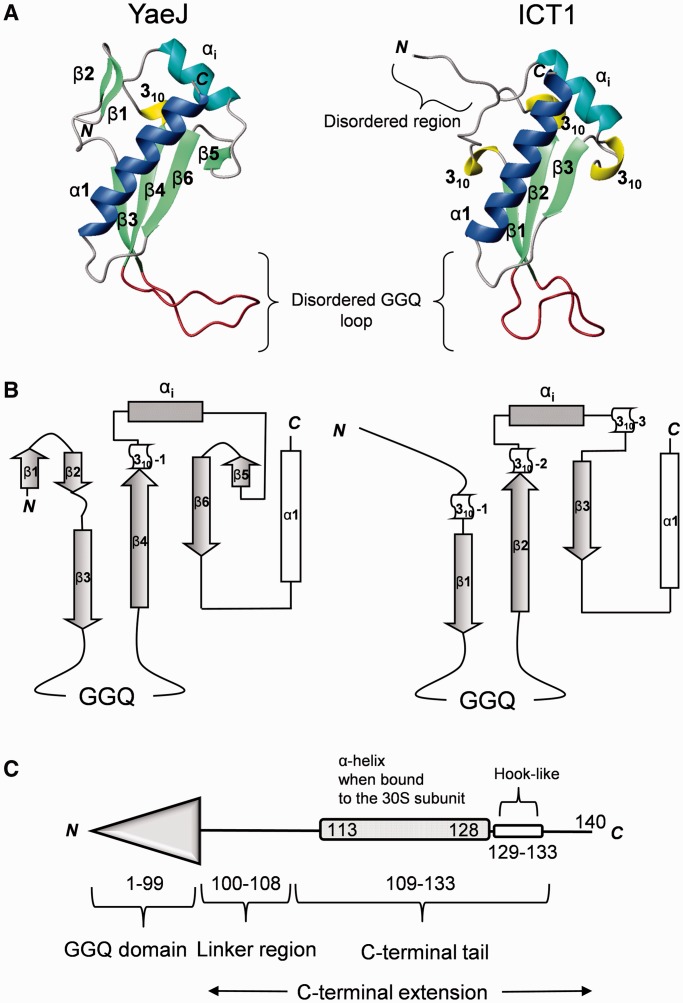
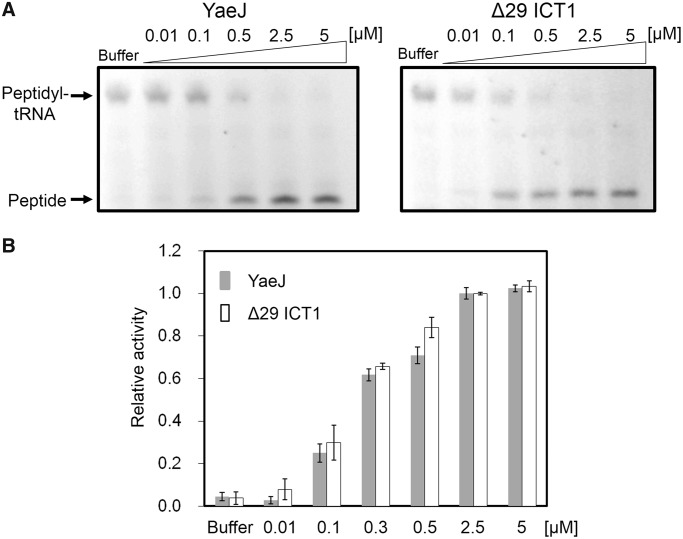
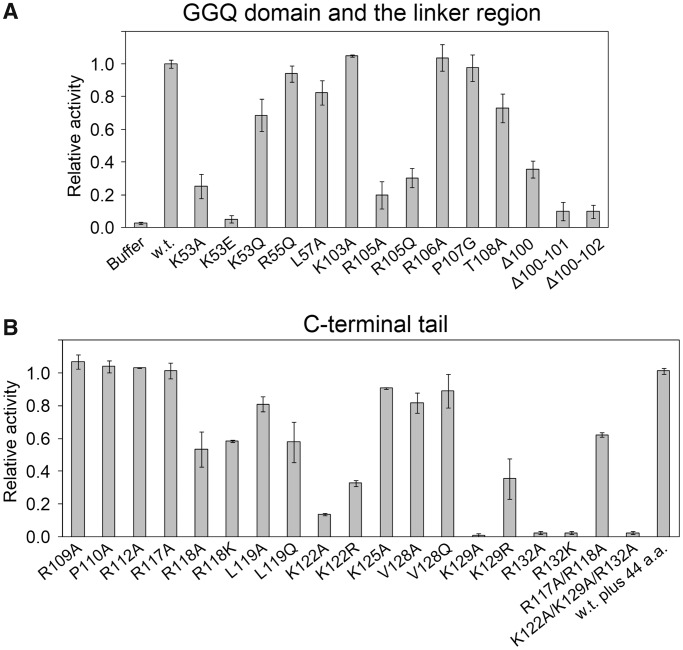
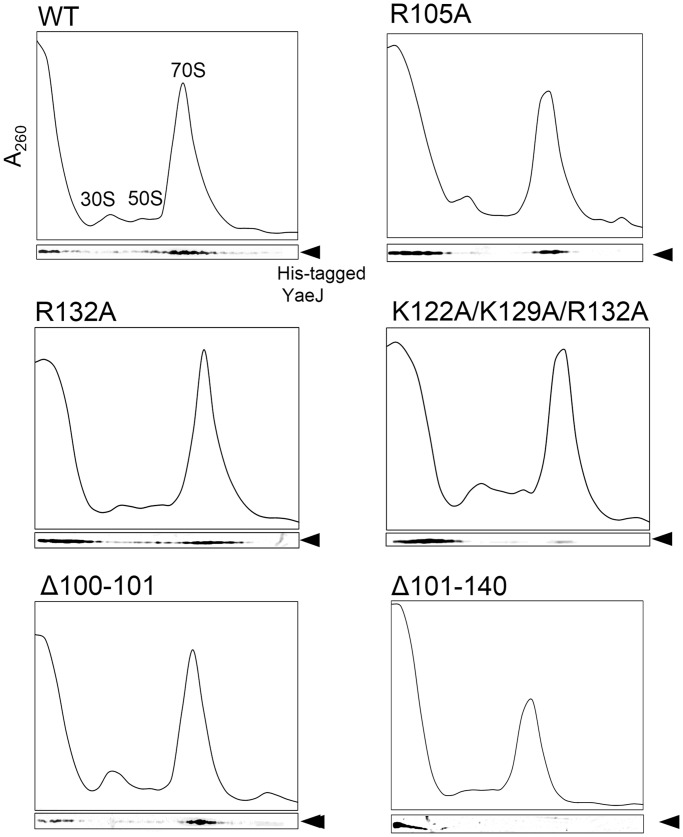
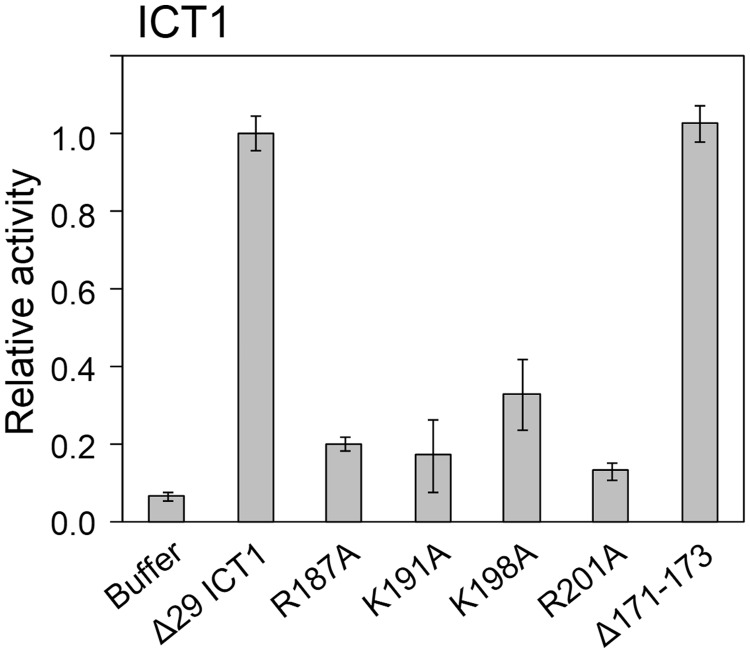
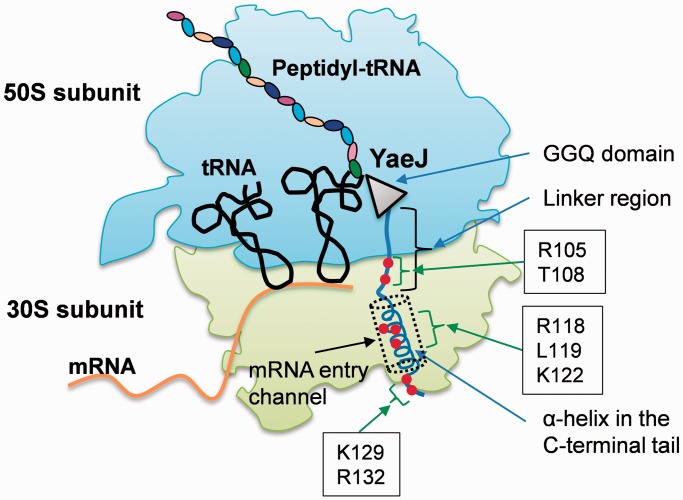
The YaeJ protein is a codon-independent release factor with peptidyl-tRNA hydrolysis (PTH) activity, and functions as a stalled-ribosome rescue factor in Escherichia coli. To identify residues required for YaeJ function, we performed mutational analysis for in vitro PTH activity towards rescue of ribosomes stalled on a non-stop mRNA, and for ribosome-binding efficiency. We focused on residues conserved among bacterial YaeJ proteins. Additionally, we determined the solution structure of the GGQ domain of YaeJ from E. coli using nuclear magnetic resonance spectroscopy. YaeJ and a human homolog, ICT1, had similar levels of PTH activity, despite various differences in sequence and structure. While no YaeJ-specific residues important for PTH activity occur in the structured GGQ domain, Arg118, Leu119, Lys122, Lys129 and Arg132 in the following C-terminal extension were required for PTH activity. All of these residues are completely conserved among bacteria. The equivalent residues were also found in the C-terminal extension of ICT1, allowing an appropriate sequence alignment between YaeJ and ICT1 proteins from various species. Single amino acid substitutions for each of these residues significantly decreased ribosome-binding efficiency. These biochemical findings provide clues to understanding how YaeJ enters the A-site of stalled ribosomes.
Figures
References
-
- Moore SD, Sauer RT. The tmRNA system for translational surveillance and ribosome rescue. Annu. Rev. Biochem. 2007;76:101–124. - PubMed
-
- Keiler KC, Waller PR, Sauer RT. Role of a peptide tagging system in degradation of proteins synthesized from damaged messenger RNA. Science. 1996;271:990–993. - PubMed
-
- Himeno H, Sato M, Tadaki T, Fukushima M, Ushida C, Muto A. In vitro trans translation mediated by alanine-charged 10Sa RNA. J. Mol. Biol. 1997;268:803–808. - PubMed
-
- Karzai AW, Roche ED, Sauer RT. The SsrA-SmpB system for protein tagging, directed degradation and ribosome rescue. Nat. Struct. Biol. 2000;7:449–455. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
