

Competing dopamine neurons drive oviposition choice for ethanol in Drosophila
- PMID: 24324162
- PMCID: PMC3876210
- DOI: 10.1073/pnas.1320208110
Competing dopamine neurons drive oviposition choice for ethanol in Drosophila
Abstract
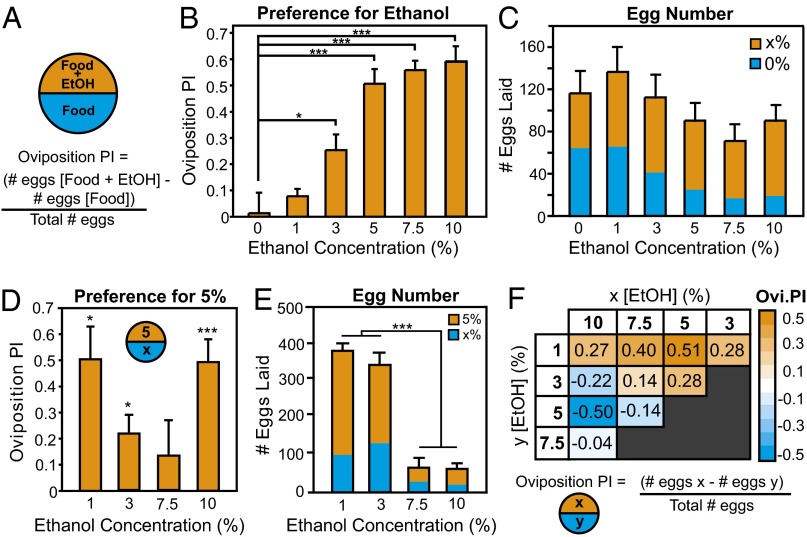
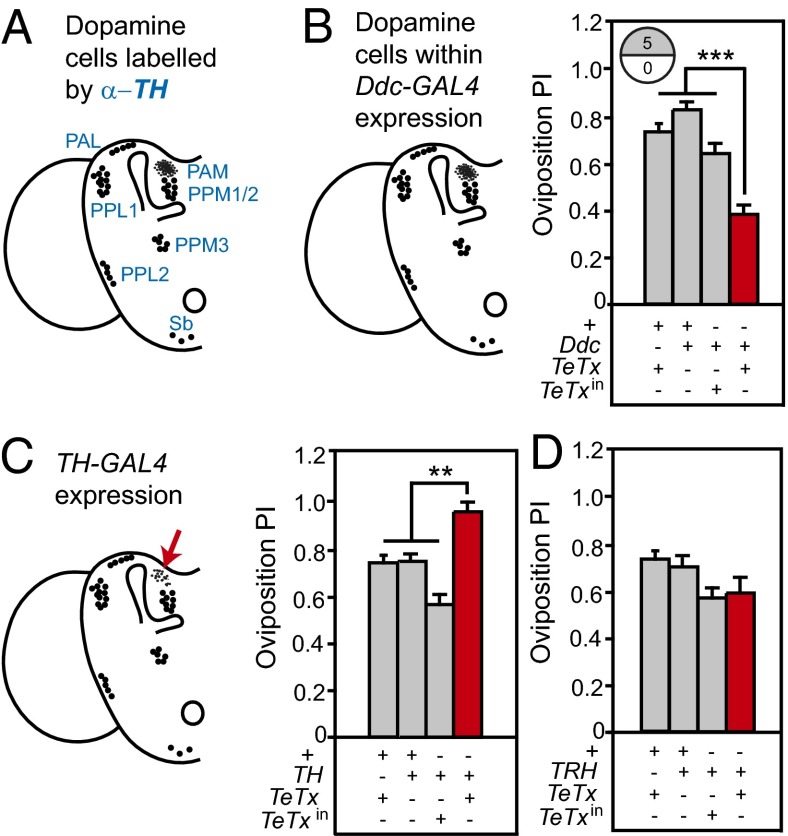
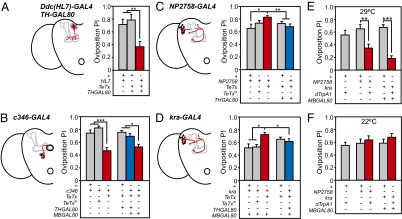
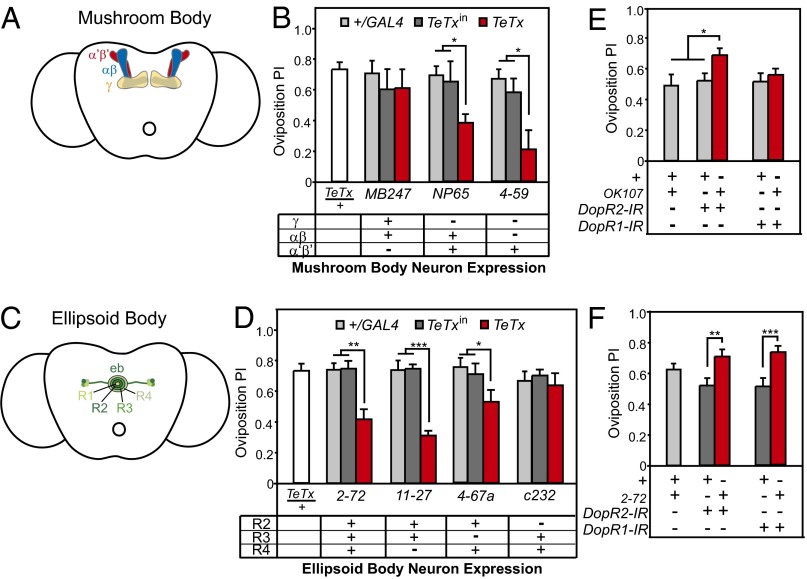
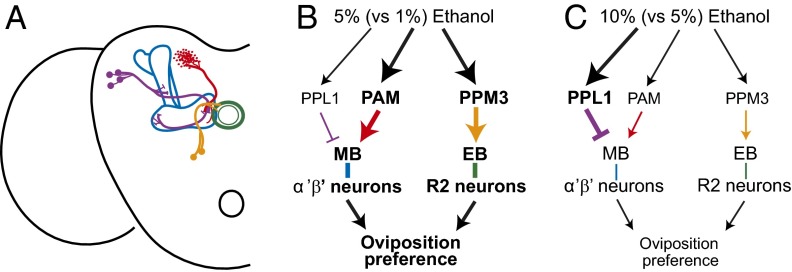
The neural circuits that mediate behavioral choice evaluate and integrate information from the environment with internal demands and then initiate a behavioral response. Even circuits that support simple decisions remain poorly understood. In Drosophila melanogaster, oviposition on a substrate containing ethanol enhances fitness; however, little is known about the neural mechanisms mediating this important choice behavior. Here, we characterize the neural modulation of this simple choice and show that distinct subsets of dopaminergic neurons compete to either enhance or inhibit egg-laying preference for ethanol-containing food. Moreover, activity in α'β' neurons of the mushroom body and a subset of ellipsoid body ring neurons (R2) is required for this choice. We propose a model where competing dopaminergic systems modulate oviposition preference to adjust to changes in natural oviposition substrates.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ashburner M. Speculations on the subject of alcohol dehydrogenase and its properties in Drosophila and other flies. Bioessays. 1998;20(11):949–954. - PubMed
-
- Dudley R. Fermenting fruit and the historical ecology of ethanol ingestion: Is alcoholism in modern humans an evolutionary hangover? Addiction. 2002;97(4):381–388. - PubMed
-
- Gibson JB, Oakeshott JG. Genetics of biochemical and behavioural aspects of alcohol metabolism. Aust N Z J Med. 1981;11(2):128–131. - PubMed
-
- Hoffmann AA, Parsons PA. Olfactory response and resource utilization in Drosophila – interspecific comparison. Biolog J Linnean Soc. 1984;22:43–53.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
