

Mutational effects on stability are largely conserved during protein evolution
- PMID: 24324165
- PMCID: PMC3876214
- DOI: 10.1073/pnas.1314781111
Mutational effects on stability are largely conserved during protein evolution
Abstract
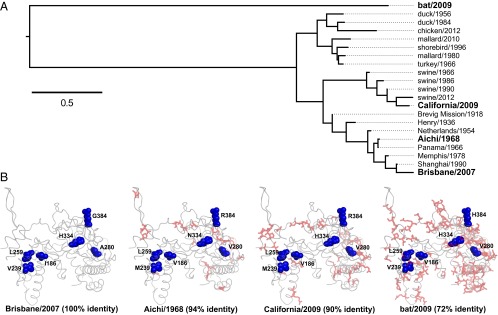
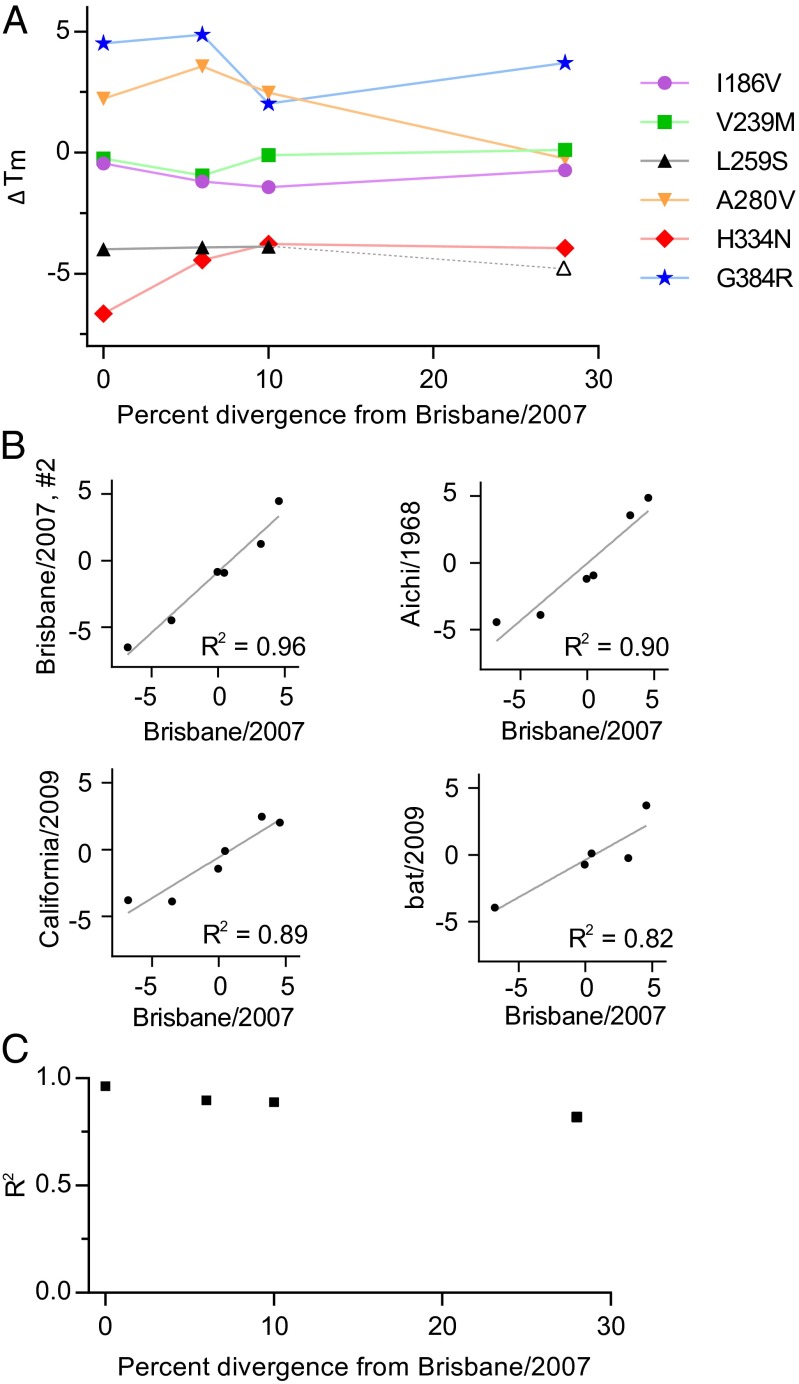
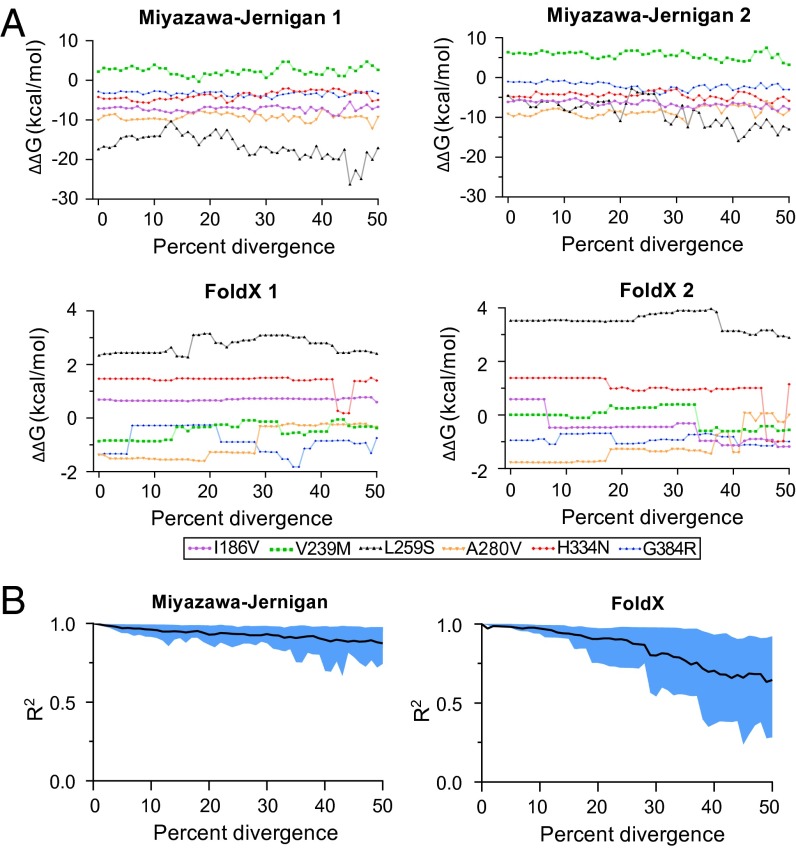
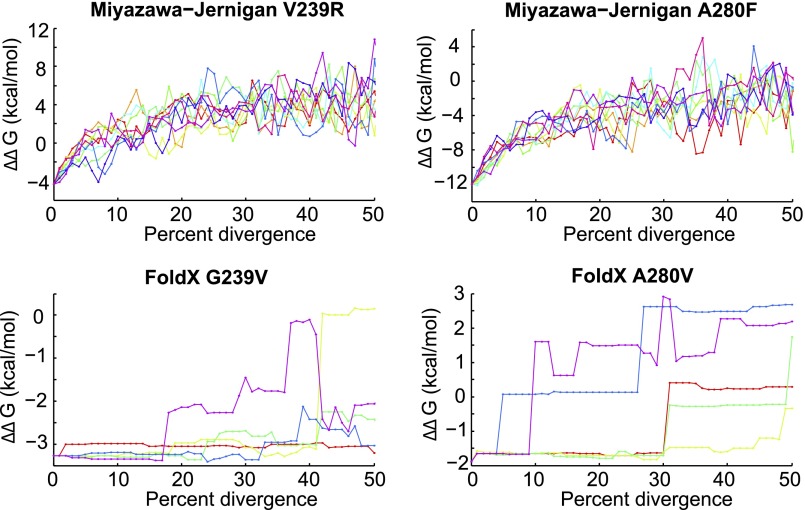
Protein stability and folding are the result of cooperative interactions among many residues, yet phylogenetic approaches assume that sites are independent. This discrepancy has engendered concerns about large evolutionary shifts in mutational effects that might confound phylogenetic approaches. Here we experimentally investigate this issue by introducing the same mutations into a set of diverged homologs of the influenza nucleoprotein and measuring the effects on stability. We find that mutational effects on stability are largely conserved across the homologs. We reach qualitatively similar conclusions when we simulate protein evolution with molecular-mechanics force fields. Our results do not mean that proteins evolve without epistasis, which can still arise even when mutational stability effects are conserved. However, our findings indicate that large evolutionary shifts in mutational effects on stability are rare, at least among homologs with similar structures and functions. We suggest that properly describing the clearly observable and highly conserved amino acid preferences at individual sites is likely to be far more important for phylogenetic analyses than accounting for rare shifts in amino acid propensities due to site covariation.
Keywords: consensus design; heterotachy; substitution models.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Strong evidence for protein epistasis, weak evidence against it.Proc Natl Acad Sci U S A. 2014 Apr 15;111(15):E1450. doi: 10.1073/pnas.1401112111. Epub 2014 Mar 20. Proc Natl Acad Sci U S A. 2014. PMID: 24706894 Free PMC article. No abstract available.
References
-
- Anfinsen CB. Principles that govern the folding of protein chains. Science. 1973;181(4096):223–230. - PubMed
-
- Rodrigue N, Philippe H, Lartillot N. Assessing site-interdependent phylogenetic models of sequence evolution. Mol Biol Evol. 2006;23(9):1762–1775. - PubMed
-
- Choi SC, Hobolth A, Robinson DM, Kishino H, Thorne JL. Quantifying the impact of protein tertiary structure on molecular evolution. Mol Biol Evol. 2007;24(8):1769–1782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
