

Canonical Wnt/β-catenin signalling is essential for optic cup formation
- PMID: 24324671
- PMCID: PMC3852023
- DOI: 10.1371/journal.pone.0081158
Canonical Wnt/β-catenin signalling is essential for optic cup formation
Abstract
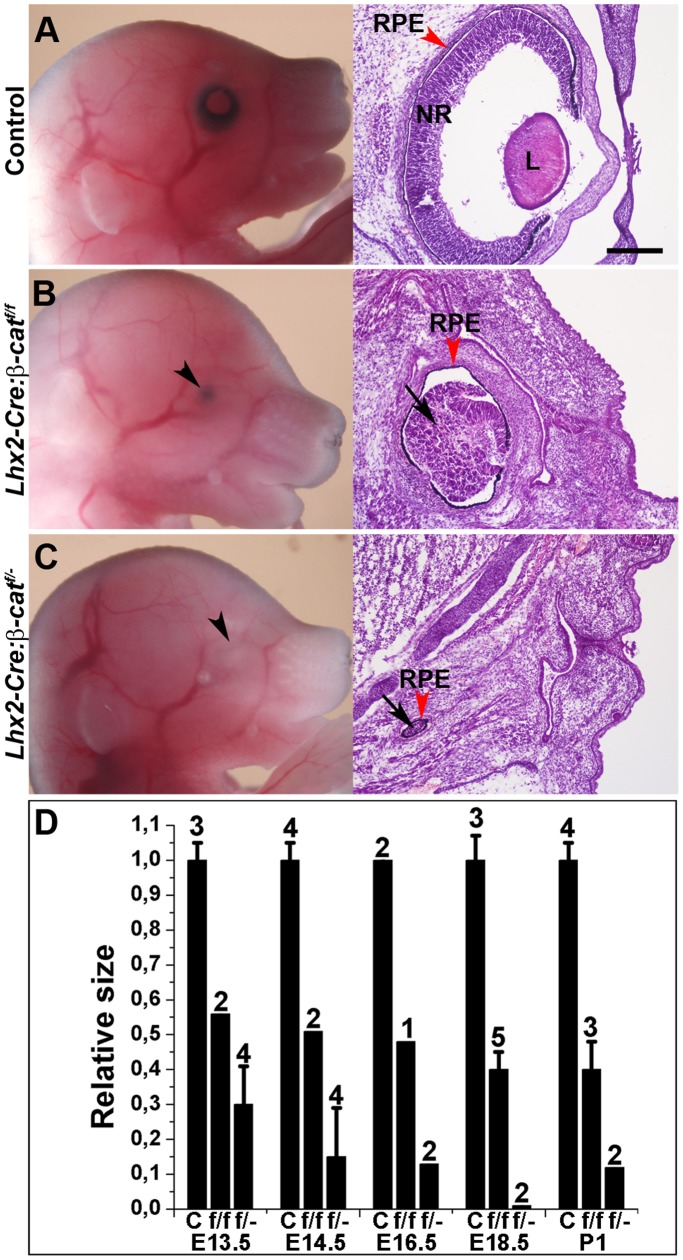
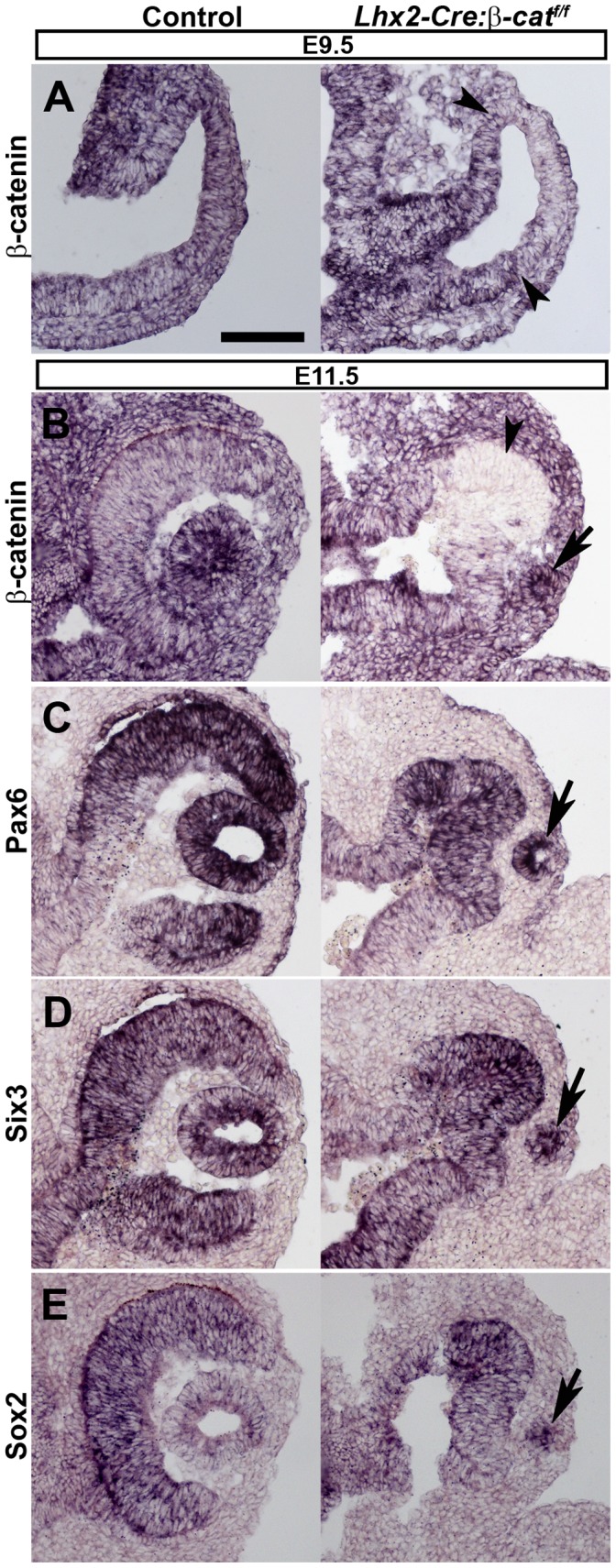
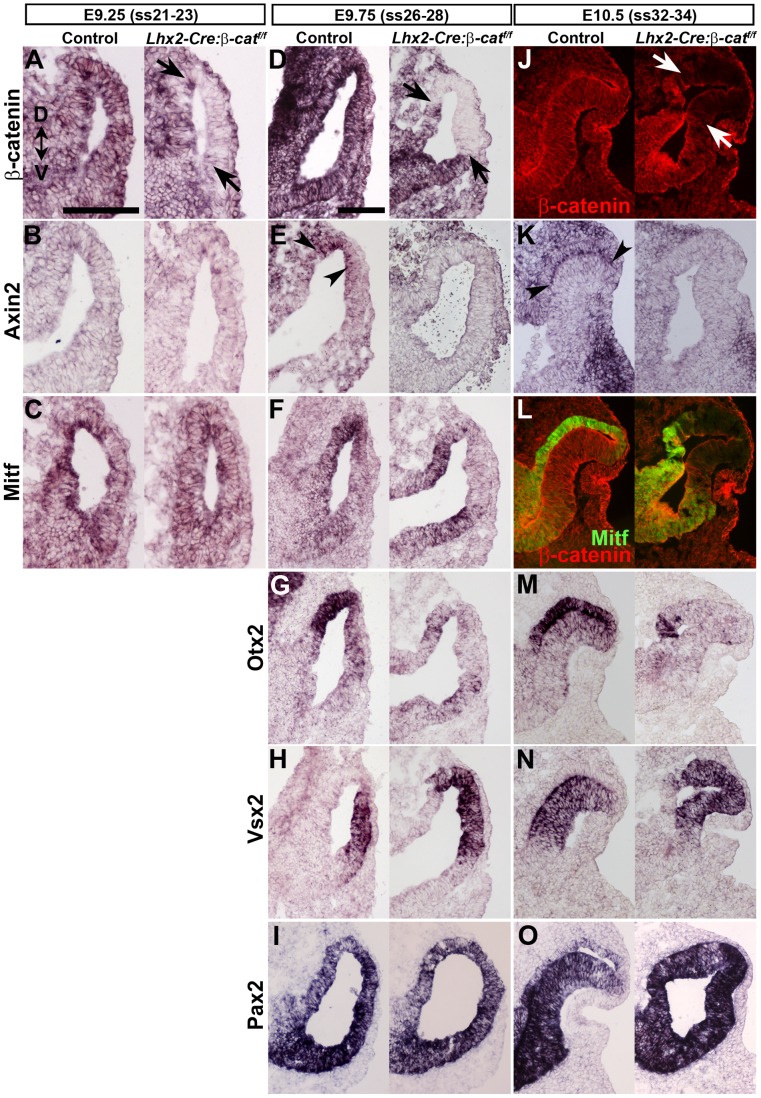
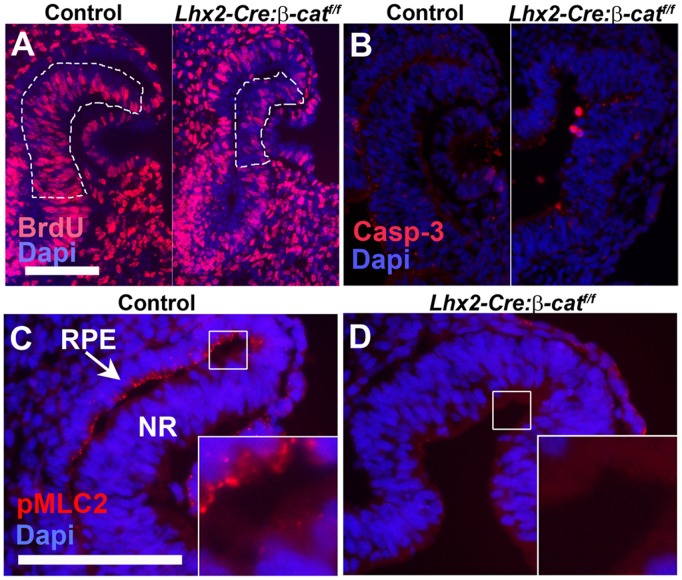
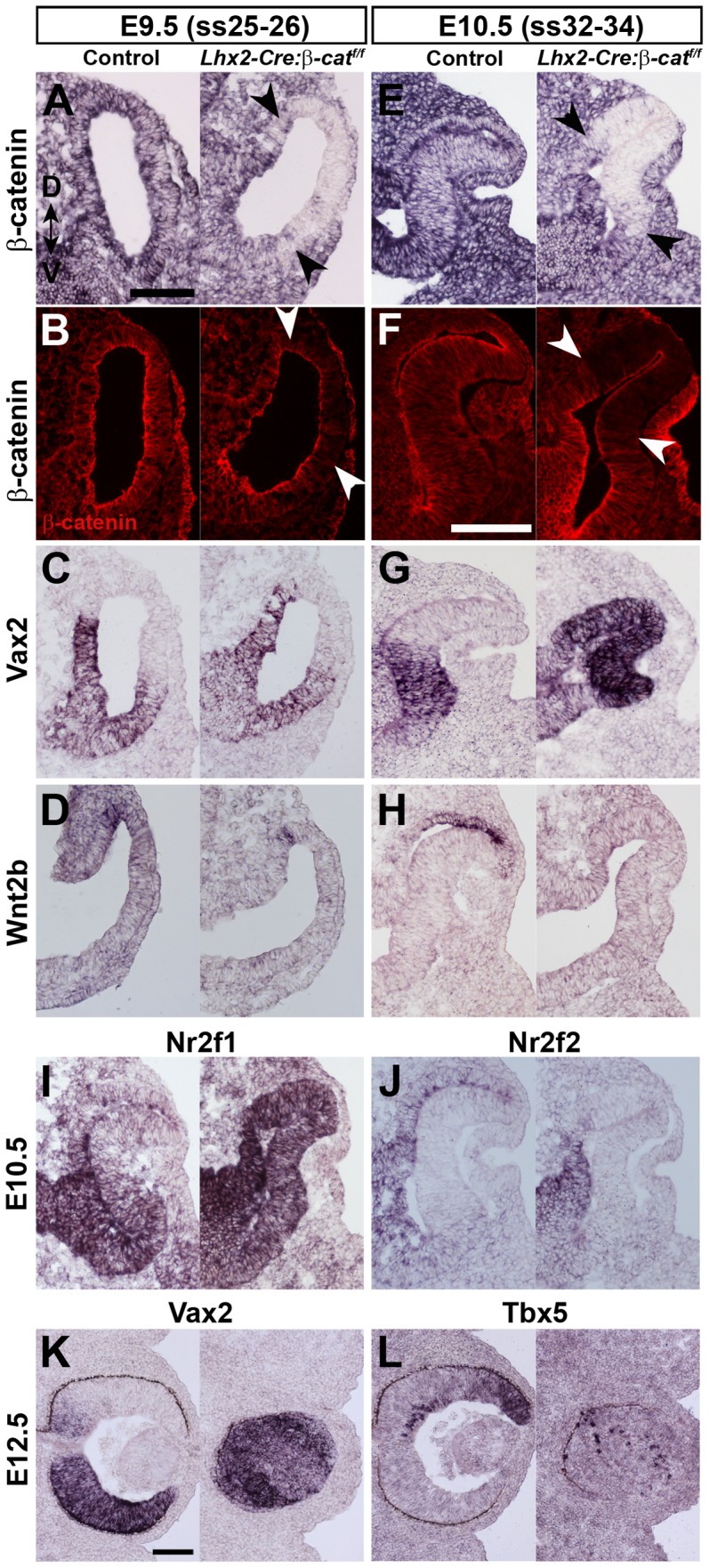
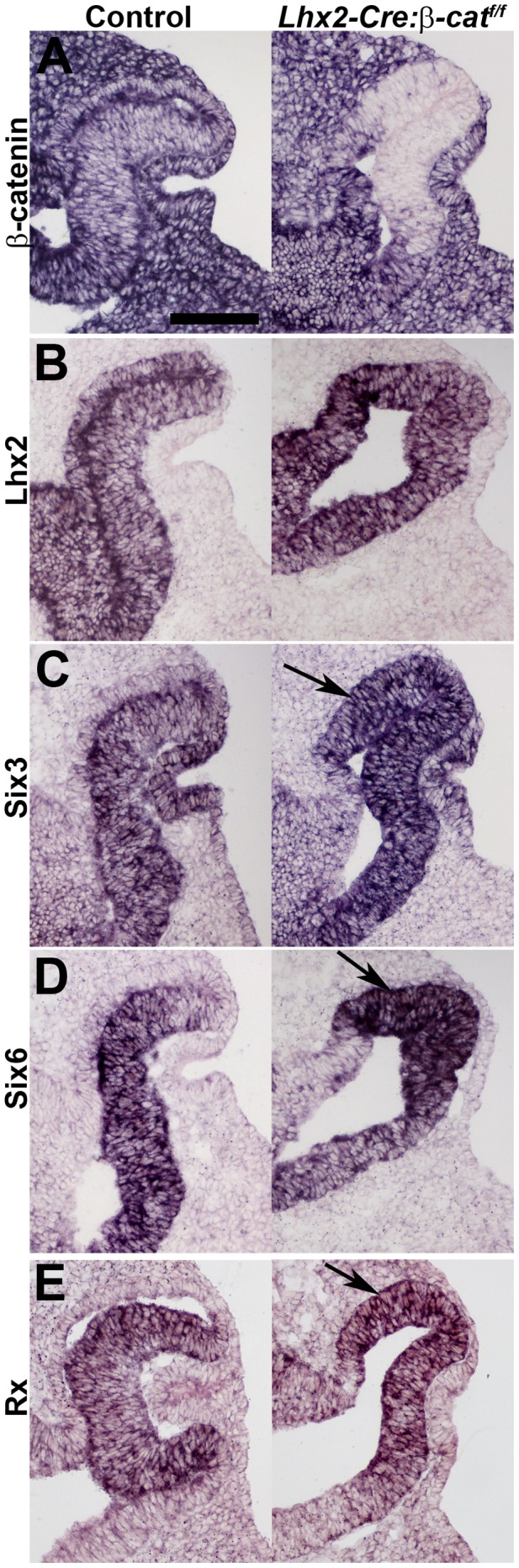
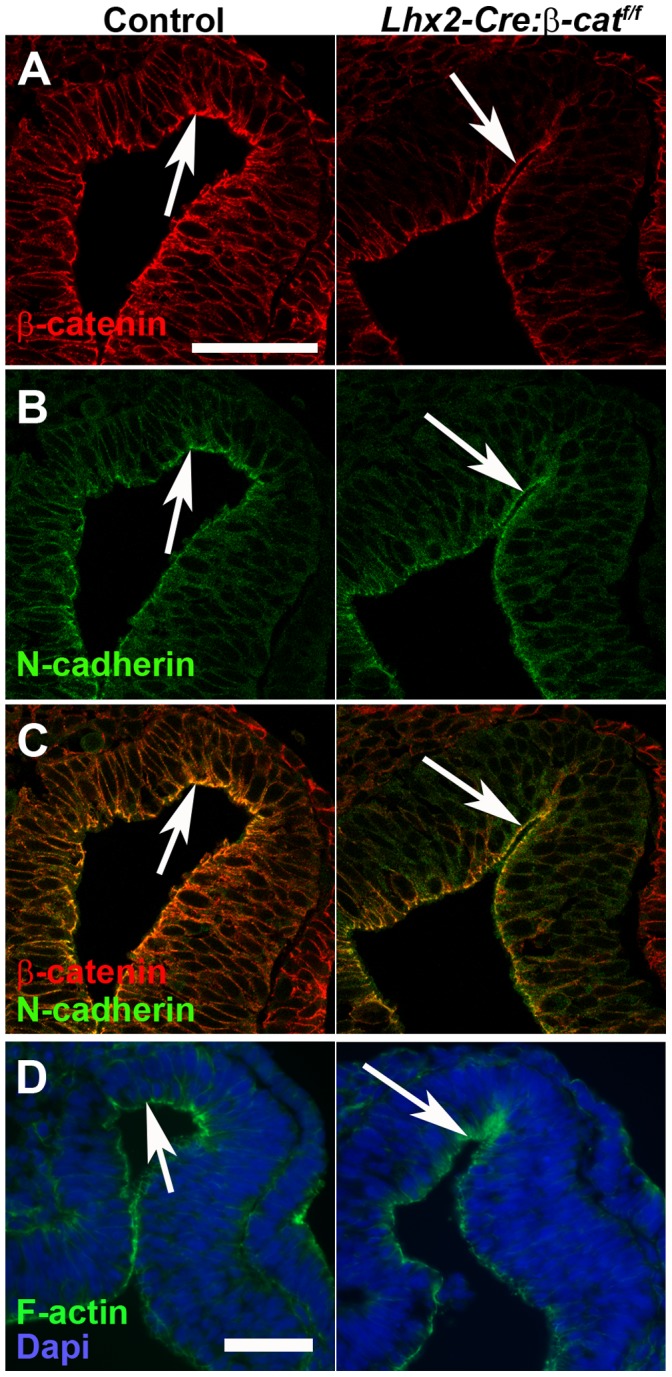
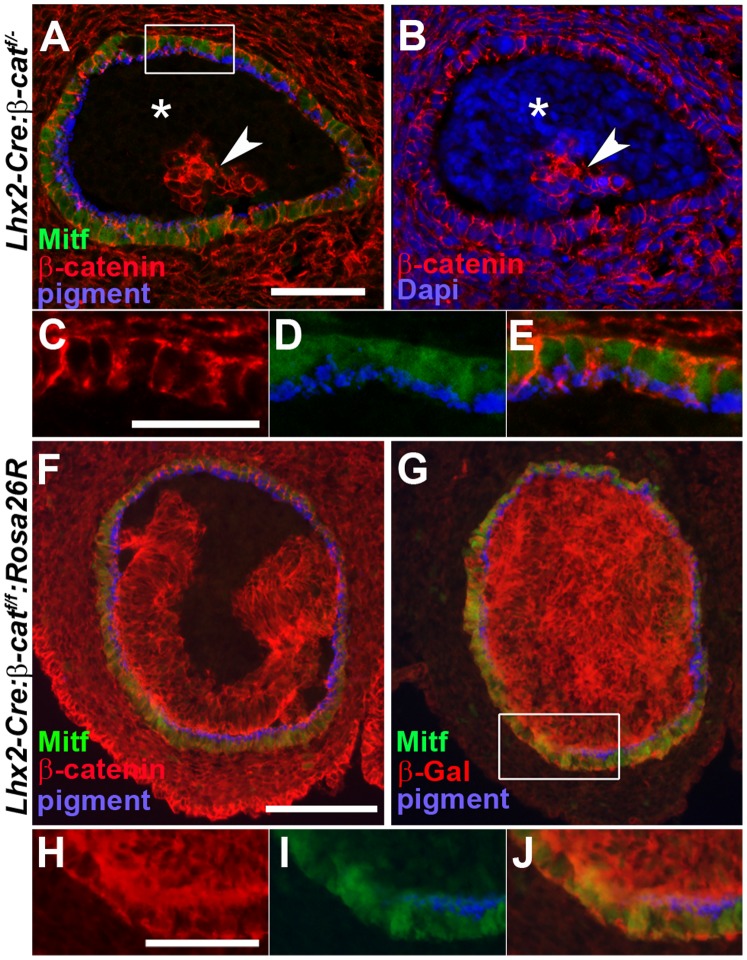
A multitude of signalling pathways are involved in the process of forming an eye. Here we demonstrate that β-catenin is essential for eye development as inactivation of β-catenin prior to cellular specification in the optic vesicle caused anophthalmia in mice. By achieving this early and tissue-specific β-catenin inactivation we find that retinal pigment epithelium (RPE) commitment was blocked and eye development was arrested prior to optic cup formation due to a loss of canonical Wnt signalling in the dorsal optic vesicle. Thus, these results show that Wnt/β-catenin signalling is required earlier and play a more central role in eye development than previous studies have indicated. In our genetic model system a few RPE cells could escape β-catenin inactivation leading to the formation of a small optic rudiment. The optic rudiment contained several neural retinal cell classes surrounded by an RPE. Unlike the RPE cells, the neural retinal cells could be β-catenin-negative revealing that differentiation of the neural retinal cell classes is β-catenin-independent. Moreover, although dorsoventral patterning is initiated in the mutant optic vesicle, the neural retinal cells in the optic rudiment displayed almost exclusively ventral identity. Thus, β-catenin is required for optic cup formation, commitment to RPE cells and maintenance of dorsal identity of the retina.
Conflict of interest statement
Figures
References
-
- Kaufman M (1994) The atlas of mouse development. San Diego: Academic Press Limited.
-
- Chow RL, Lang RA (2001) Early eye development in vertebrates. Annu Rev Cell Dev Biol 17: 255–296. - PubMed
-
- Eiraku M, Takata N, Ishibashi H, Kawada M, Sakakura E, et al. (2011) Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature 472: 51–56. - PubMed
-
- Horsford DJ, Nguyen MT, Sellar GC, Kothary R, Arnheiter H, et al. (2005) Chx10 repression of Mitf is required for the maintenance of mammalian neuroretinal identity. Development 132: 177–187. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
