

Early dynamic fate changes in haemogenic endothelium characterized at the single-cell level
- PMID: 24326267
- PMCID: PMC4362711
- DOI: 10.1038/ncomms3924
Early dynamic fate changes in haemogenic endothelium characterized at the single-cell level
Abstract
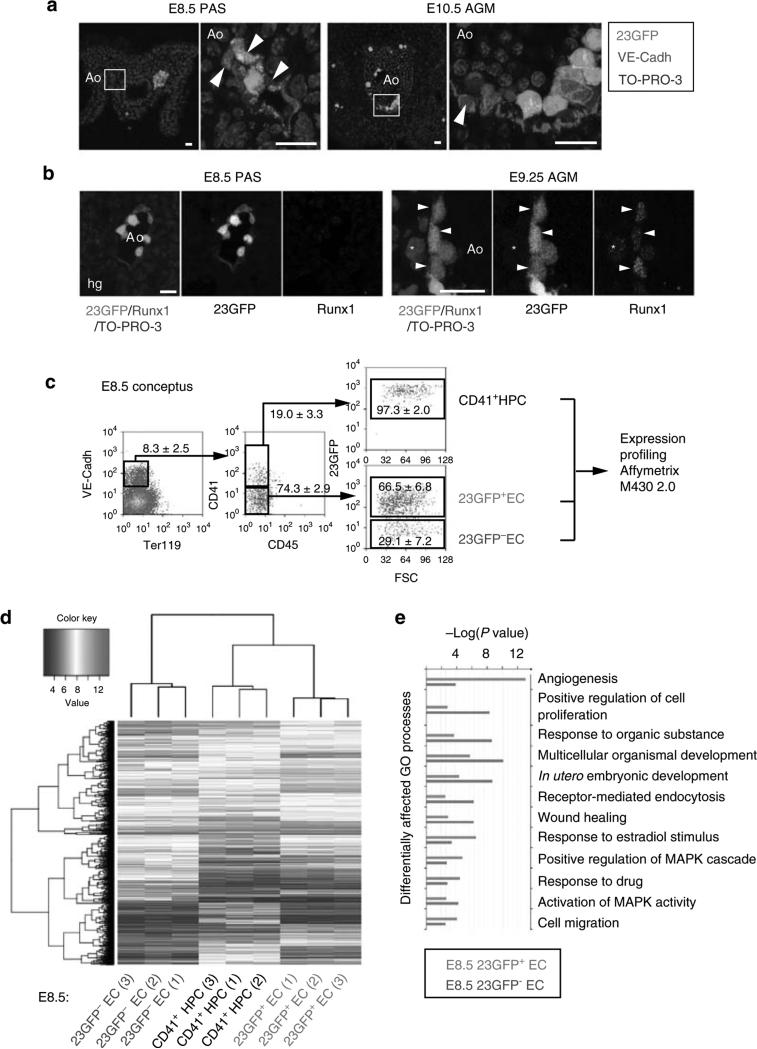
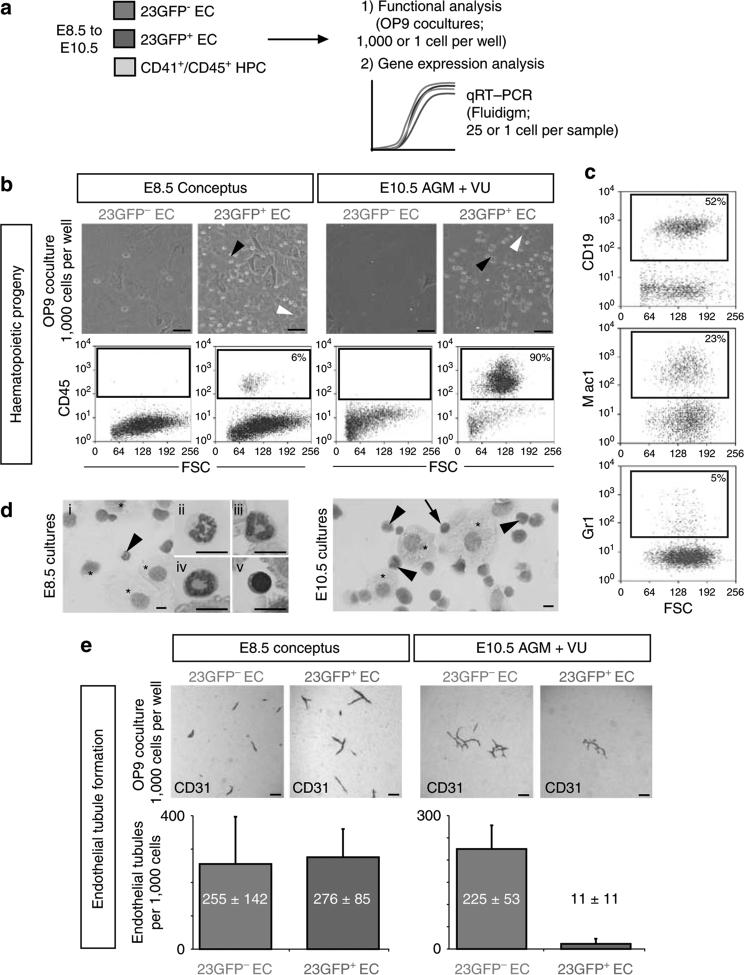
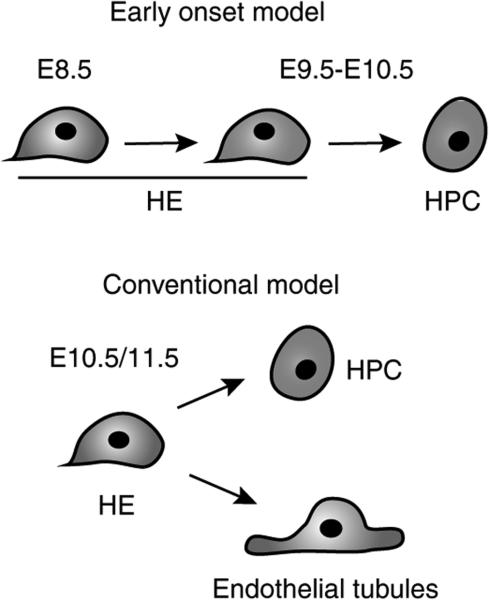
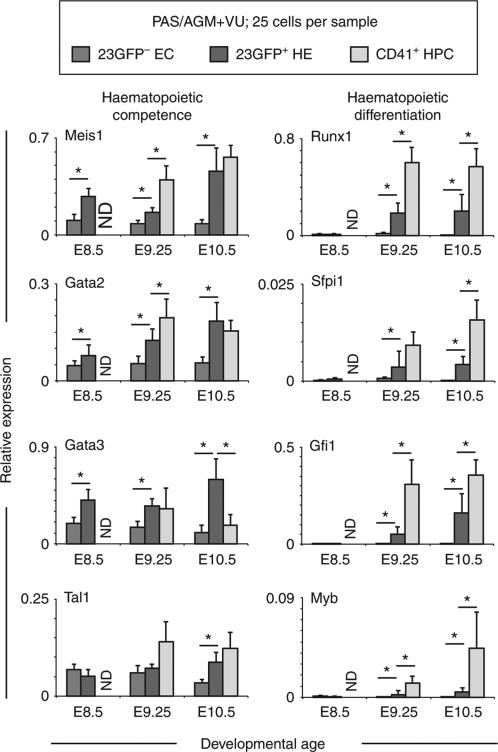
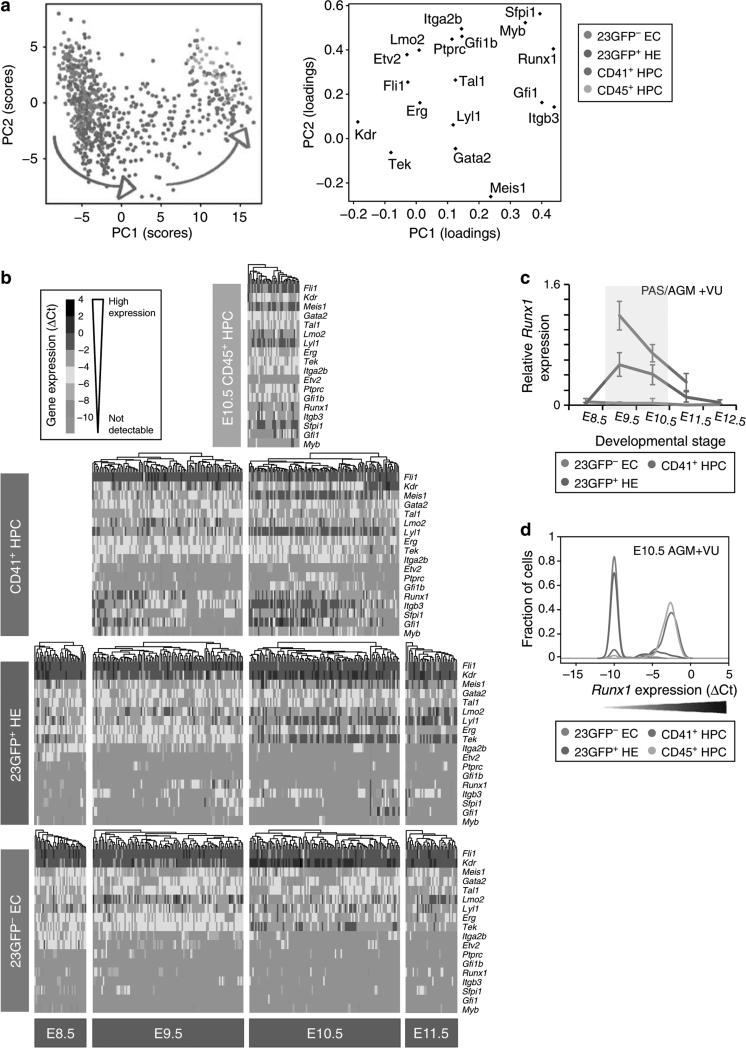
Haematopoietic stem cells (HSCs) are the founding cells of the adult haematopoietic system, born during ontogeny from a specialized subset of endothelium, the haemogenic endothelium (HE) via an endothelial-to-haematopoietic transition (EHT). Although recently imaged in real time, the underlying mechanism of EHT is still poorly understood. We have generated a Runx1 +23 enhancer-reporter transgenic mouse (23GFP) for the prospective isolation of HE throughout embryonic development. Here we perform functional analysis of over 1,800 and transcriptional analysis of 268 single 23GFP(+) HE cells to explore the onset of EHT at the single-cell level. We show that initiation of the haematopoietic programme occurs in cells still embedded in the endothelial layer, and is accompanied by a previously unrecognized early loss of endothelial potential before HSCs emerge. Our data therefore provide important insights on the timeline of early haematopoietic commitment.
Figures
References
-
- Dieterlen-Lievre F. Hematopoiesis: progenitors and their genetic program. Curr. Biol. 1998;8:R727–R730. - PubMed
-
- Jaffredo T, et al. From hemangioblast to hematopoietic stem cell: an endothelial connection? Exp. Hematol. 2005;33:1029–1040. - PubMed
-
- Medvinsky A, Rybtsov S, Taoudi S. Embryonic origin of the adult hematopoietic system: advances and questions. Development. 2011;138:1017–1031. - PubMed
-
- Antas VI, Al-Drees MA, Prudence AJ, Sugiyama D, Fraser ST. Hemogenic endothelium: a vessel for blood production. Int. J. Biochem. Cell Biol. 2012;45:692–695. - PubMed
-
- Eilken HM, Nishikawa S, Schroeder T. Continuous single-cell imaging of blood generation from haemogenic endothelium. Nature. 2009;457:896–900. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- MC_U137970202/MRC_/Medical Research Council/United Kingdom
- 079249/WT_/Wellcome Trust/United Kingdom
- G1000801H/MRC_/Medical Research Council/United Kingdom
- G0900729/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- MC_UU_12009/2/MRC_/Medical Research Council/United Kingdom
- 12765/CRUK_/Cancer Research UK/United Kingdom
- MC_U137973817/MRC_/Medical Research Council/United Kingdom
- MC_QA137913/MRC_/Medical Research Council/United Kingdom
- 12796/CRUK_/Cancer Research UK/United Kingdom
- MC_PC_12009/MRC_/Medical Research Council/United Kingdom
- R01 HL063169/HL/NHLBI NIH HHS/United States
- 100140/WT_/Wellcome Trust/United Kingdom
- G1000801G/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
