

Fitness benefits in fluoroquinolone-resistant Salmonella Typhi in the absence of antimicrobial pressure
- PMID: 24327559
- PMCID: PMC3857714
- DOI: 10.7554/eLife.01229
Fitness benefits in fluoroquinolone-resistant Salmonella Typhi in the absence of antimicrobial pressure
Abstract
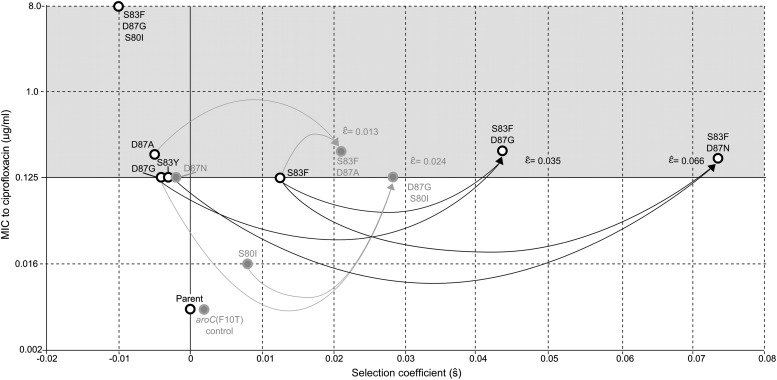
Fluoroquinolones (FQ) are the recommended antimicrobial treatment for typhoid, a severe systemic infection caused by the bacterium Salmonella enterica serovar Typhi. FQ-resistance mutations in S. Typhi have become common, hindering treatment and control efforts. Using in vitro competition experiments, we assayed the fitness of eleven isogenic S. Typhi strains with resistance mutations in the FQ target genes, gyrA and parC. In the absence of antimicrobial pressure, 6 out of 11 mutants carried a selective advantage over the antimicrobial-sensitive parent strain, indicating that FQ resistance in S. Typhi is not typically associated with fitness costs. Double-mutants exhibited higher than expected fitness as a result of synergistic epistasis, signifying that epistasis may be a critical factor in the evolution and molecular epidemiology of S. Typhi. Our findings have important implications for the management of drug-resistant S. Typhi, suggesting that FQ-resistant strains would be naturally maintained even if fluoroquinolone use were reduced. DOI: http://dx.doi.org/10.7554/eLife.01229.001.
Keywords: Salmonella; epistasis; fitness cost; fluoroquinolone; typhoid.
Conflict of interest statement
JF: Jeremy Farrar is Director of the Wellcome Trust, one of the three founding funders of
The other authors declare that no competing interests exist.
Figures





Comment in
-
Fitness gains hamper efforts to tackle drug resistance.Elife. 2013 Dec 10;2:e01809. doi: 10.7554/eLife.01809. Elife. 2013. PMID: 24327563 Free PMC article.
-
Microbial genetics: Fitness from drug resistance.Nat Rev Genet. 2014 Feb;15(2):64. doi: 10.1038/nrg3658. Epub 2014 Jan 3. Nat Rev Genet. 2014. PMID: 24384548 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
