

Effect of Cyp27A1 gene dosage on atherosclerosis development in ApoE-knockout mice
- PMID: 24327605
- PMCID: PMC4046167
- DOI: 10.1096/fj.13-233791
Effect of Cyp27A1 gene dosage on atherosclerosis development in ApoE-knockout mice
Abstract
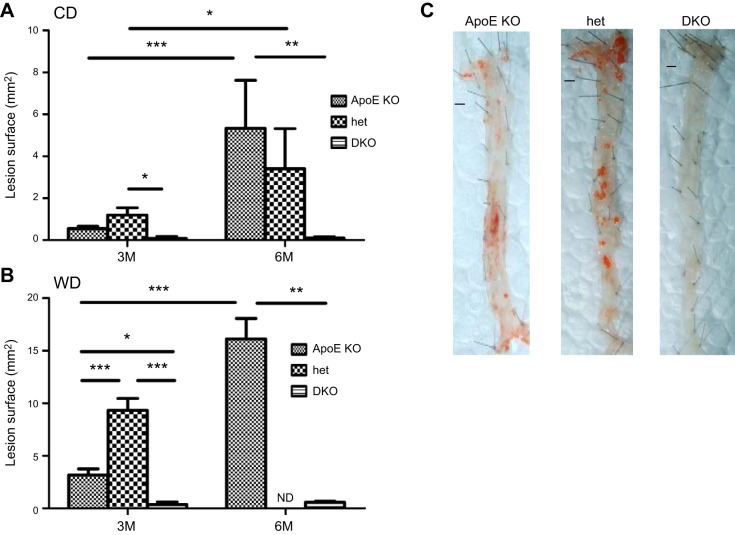
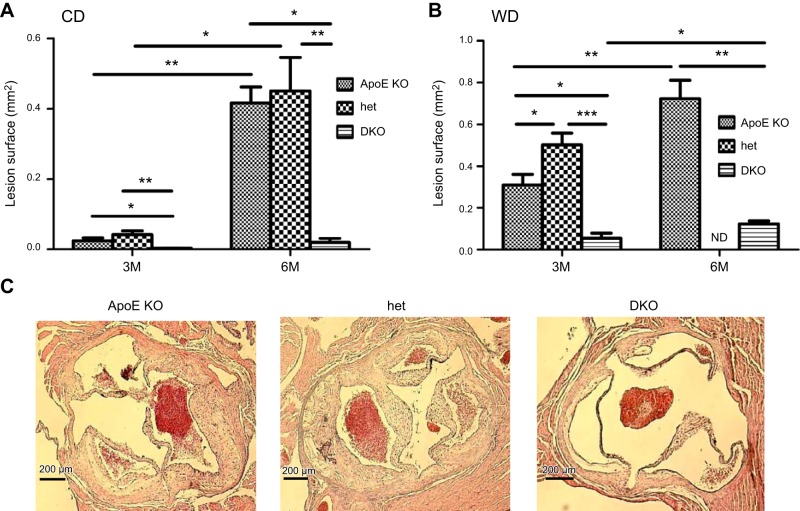
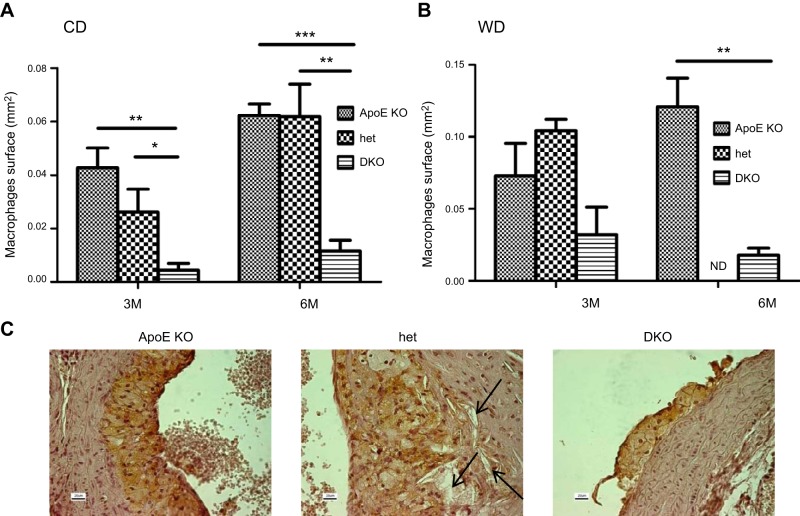
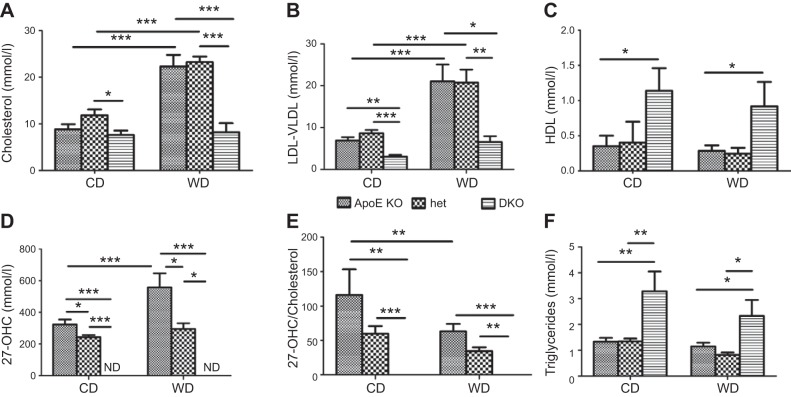
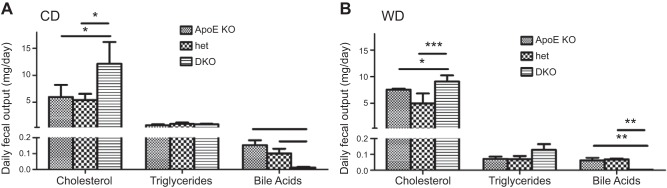
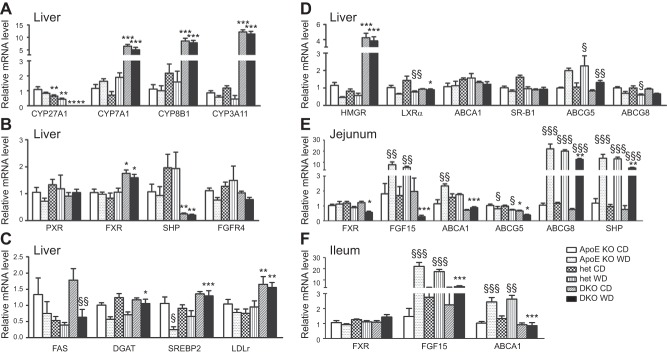
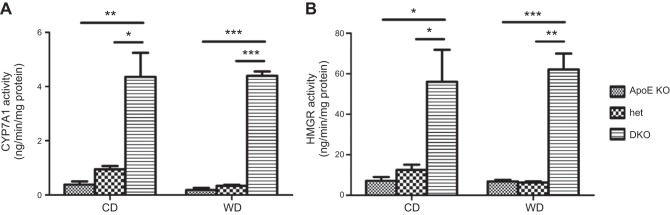
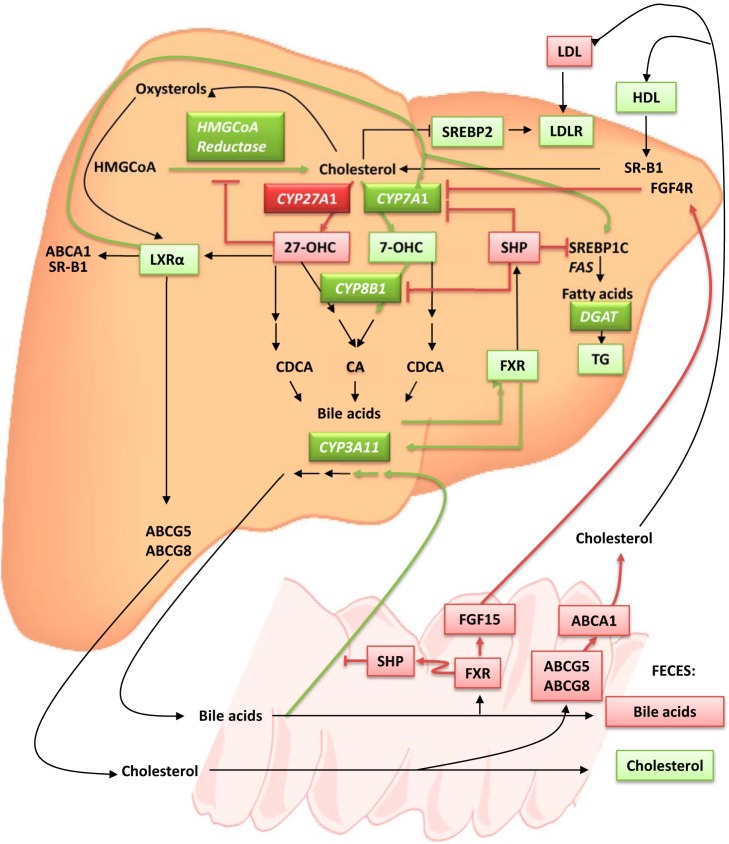
In humans, sterol 27-hydroxylase (CYP27A1) deficiency leads to cholesterol deposition in tendons and vasculature. Thus, in addition to its role in bile acid synthesis, where it converts cholesterol to 27-hydroxycholesterol (27-OHC), CYP27A1 may also be atheroprotective. Cyp27A1-deficient (Cyp27A1(-/-)) mice were crossed with apolipoprotein E (apoE)-deficient mice. Cyp27A1(+/+)/apoE(-/-) [ApoE-knockout (KO)], Cyp27A1(+/-)/apoE(-/-) heterozygous (het), and Cyp27A1(-/-)/apoE(-/-) [double-knockout (DKO)] mice were challenged with a Western diet (WD) for 3 and 6 mo. ApoE-KO mice fed a chow diet or a WD were used as the control. The severity of atherosclerosis in DKO mice was reduced 10-fold. Compared with the control, the DKO mice had no 27-OHC, total plasma cholesterol and low-density lipoprotein and very low density lipoprotein (LDL/VLDL) concentrations were reduced 2-fold, and HDL was elevated 2-fold. Expression of hepatic CYP7A1, CYP3A, and CYP8B1 were 5- to 10-fold higher. 3-Hydroxy-3-methyl-glutaryl-CoA reductase (HMGR) activity increased 4-fold. Fecal cholesterol was increased. In contrast, het mice fed a WD developed accelerated atherosclerosis and severe skin lesions, possibly because of reduced reverse cholesterol transport due to diminished 27-OHC production. CYP27A1 activity is involved in the control of cholesterol homeostasis and development of atherosclerosis with a distinct gene dose-dependent effect.
Keywords: 27-hydroxycholesterol; CYP3A11; LDL/VLDL fraction.
Figures
References
-
- Norlin M., von Bahr S., Bjorkhem I., Wikvall K. (2003) On the substrate specificity of human CYP27A1: implications for bile acid and cholestanol formation. J. Lipid Res. 44, 1515–1522 - PubMed
-
- Bjorkhem I. (1992) Mechanism of degradation of the steroid side chain in the formation of bile acids. J. Lipid Res. 33, 455–471 - PubMed
-
- Russell D. W., Setchell K. D. (1992) Bile acid biosynthesis. Biochemistry 31, 4737–4749 - PubMed
-
- Escher G., Krozowski Z., Croft K. D., Sviridov D. (2003) Expression of sterol 27-hydroxylase (CYP27A1) enhances cholesterol efflux. J. Biol. Chem. 278, 11015–11019 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
