

doi: 10.1186/1741-7007-11-119.
Why are bacteria different from eukaryotes?
Affiliations
- PMID: 24330667
- PMCID: PMC3874686
- DOI: 10.1186/1741-7007-11-119
Item in Clipboard
Why are bacteria different from eukaryotes?
BMC Biol.
.
No abstract available
Figures

Julie Theriot.
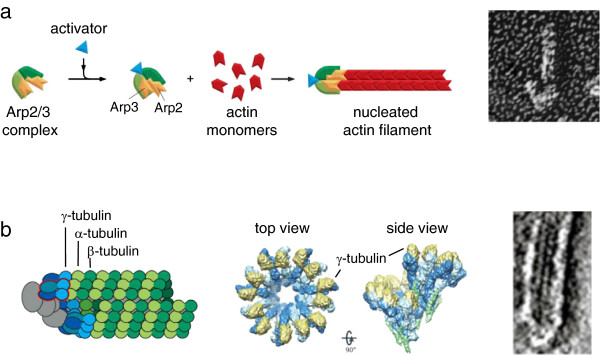
Cytoskeletal filament nucleation by modified subunits. (a) Nucleation of actin filaments by the Arp2/3 complex. Left: diagram of Arp2/3 complex before and after activation, showing rearrangement of actin-like subunits leading to templated filament growth (Copyright 2008 from Molecular Biology of the Cell, 5th edition by Alberts et al. Reproduced by permission of Garland Science/Taylor & Francis LLC [41]). Right: electron micrograph showing the appearance of an actin filament nucleated by Arp2/3 (at the bottom) (from Proc Natl Acad Sci U S A[35]). (b) Nucleation of microtubules by the γ-tubulin ring complex. Left: diagram of microtubule templated from a ring complex (Copyright 2008 from Molecular Biology of the Cell, 5th edition by Alberts et al. Reproduced by permission of Garland Science/Taylor & Francis LLC [41]). Middle, structure of the ring complex by cryo-electron microscopy, showing how the γ-tubulins are held in the proper configuration to imitate a microtubule plus end (reprinted by permission from Macmillan Publishers Ltd: Nat Rev Mol Cell Biol12:709–721, copyright 2011 [38]). Right, electron micrograph of the end of a microtubule nucleated from a ring complex (reprinted by permission from Macmillan Publishers Ltd: Nat Cell Biol2:365–370, copyright 2000 [42]).
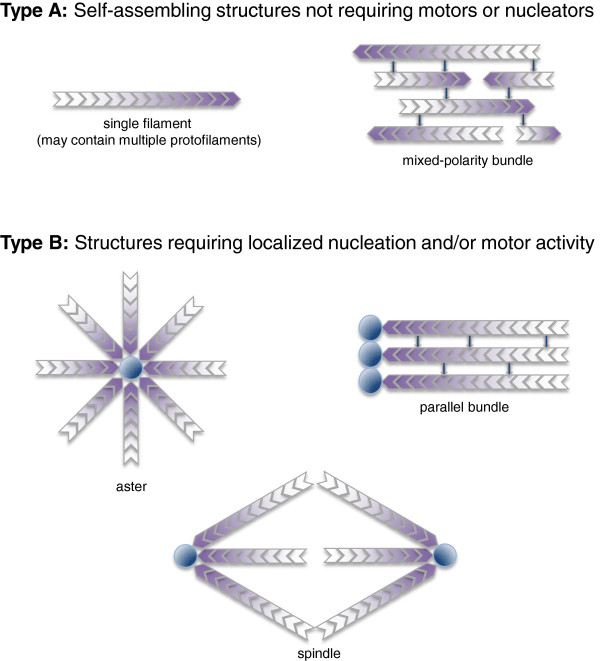
Types of cytoskeletal filament arrays. Type A: simple filament arrays that can self-assemble in the absence of spatially regulated nucleators or molecular motor protiens. Shading indicates the orientation of filament polarity. Type B: complex filament arrays that require either nucleation or motor protein activity, or both. Dark circles represent nucleators.
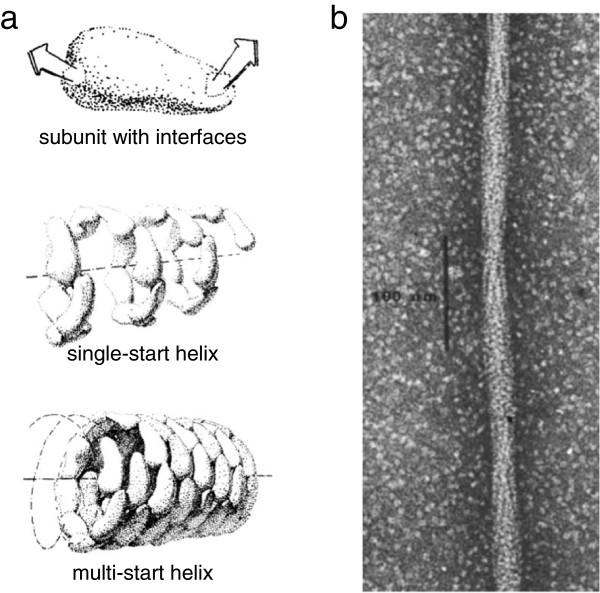
Helical protein filaments formed by self-assembly. (a) General scheme for protein self-assembly into helices. For any globular protein of arbitrary shape, as shown at the top, considered as interacting with a second copy of itself in all possible orientations, there will be some pair of surface patches that result in optimal binding energy. It is highly unlikely that those two interface patches will happen to reflect any specific geometrical symmetry. When many copies of the same subunit self-associate by binding to one another through these surface interactions, a one-start helix with a single protofilament is the default structure formed, as shown in the middle. At bottom, if weaker interactions can also form laterally between subunits, multi-start helices may be stabilized (adapted with permission from the Royal Society of Chemistry [62]). (b) Electron micrograph showing a single filament of sickle-cell hemoglobin (HbS) (reprinted by permission from Macmillan Publishers Ltd: Nature272:506–510, copyright 1978 [63]).
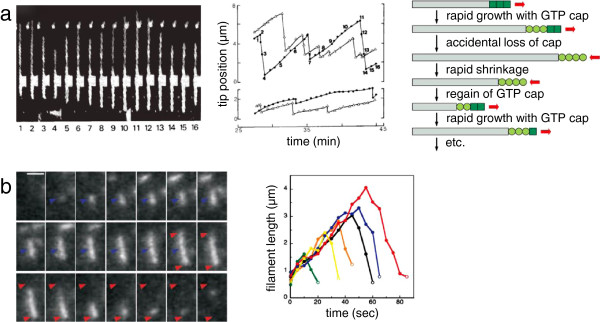
Dynamic instability of cytoskeletal filaments from eukaryotes and bacteria. (a) Dynamic instability of eukaryotic microtubules. Left: direct observation using dark-field microscopy of a microtubule undergoing dynamic instability. Middle: graph showing position of plus ends (top) and minus ends (bottom) for two dynamically unstable microtubules, with repeated cycles of growth and shrinkage. Numbered points correspond to individual video frames as labeled on the left (reprinted by permission from Macmillan Publishers Ltd: Nature321:605–607, copyright 1986 [69]). Right, schematic diagram showing the connection between nucleotide hydrolysis and dynamic instability (Copyright 2008 from Molecular Biology of the Cell, 5th edition by Alberts et al. Reproduced by permission of Garland Science/Taylor & Francis LLC [41]). (b) Dynamic instability of bacterial ParM filaments. Left: fluorescence time-lapse images of a single ParM filament over time. Blue arrowhead shows position of initial filament appearance; red arrowheads mark the most extreme positions of the two tips. Video frames are separated in time by 5 s; scale bar is 2 μm. Right, traces of filament length over time for six different ParM filaments, showing a phase of growth followed by catastrophic shrinking. (From Garner EC, Campbell CS, Mullins RD: Dynamic instability in a DNA-segregating prokaryotic actin homolog. Science 2004, 306:1021–1025. Reprinted with permission from AAAS [54].)
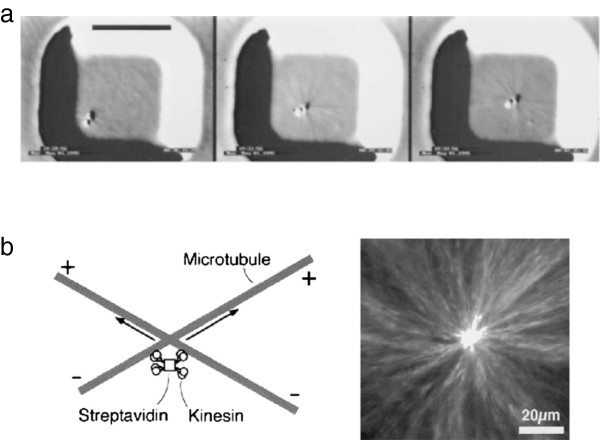
Self-centering activity of dynamic microtubule arrays. (a) Self-centering by nucleation and dynamic instability, for a microtubule-nucleating bead. Images are separated by 3 minutes, scale bar is 10 μm (From Proc Natl Acad Sci U S A[76]). (b) Self-centering by motors. Left: diagram of crosslinked motors reorienting microtubules. Right: fluorescence image of an aster formed in a microwell by this mechanism (reprinted by permission from Macmillan Publishers Ltd: Nature389:305–308, copyright 1997 [79]).
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
