

Genome-wide analyses implicate 33 loci in heritable dog osteosarcoma, including regulatory variants near CDKN2A/B
- PMID: 24330828
- PMCID: PMC4053774
- DOI: 10.1186/gb-2013-14-12-r132
Genome-wide analyses implicate 33 loci in heritable dog osteosarcoma, including regulatory variants near CDKN2A/B
Abstract
Background: Canine osteosarcoma is clinically nearly identical to the human disease, but is common and highly heritable, making genetic dissection feasible.
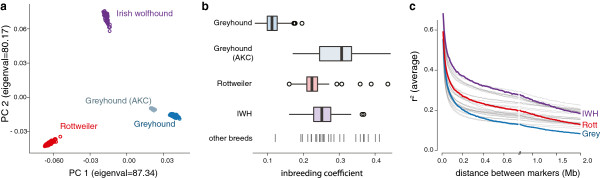
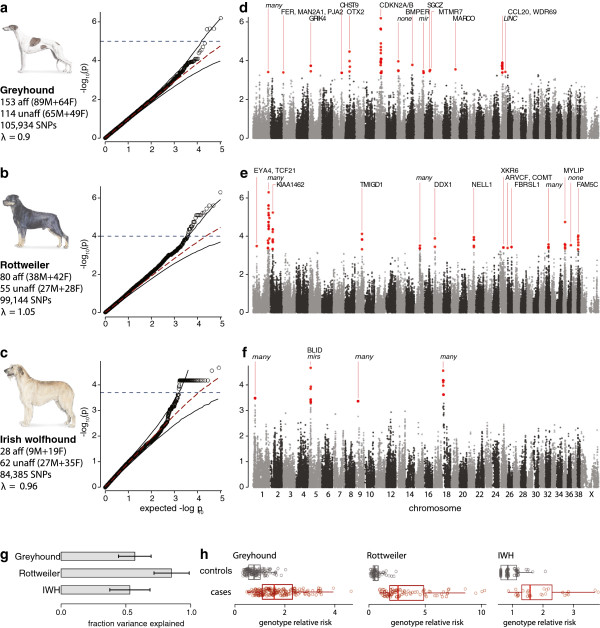
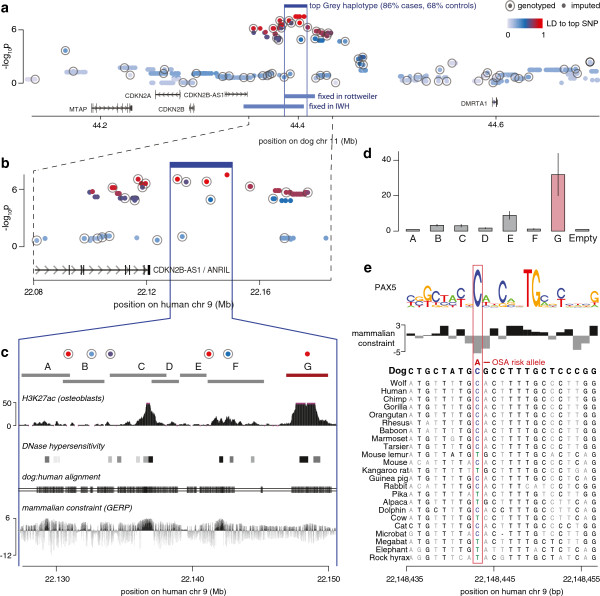
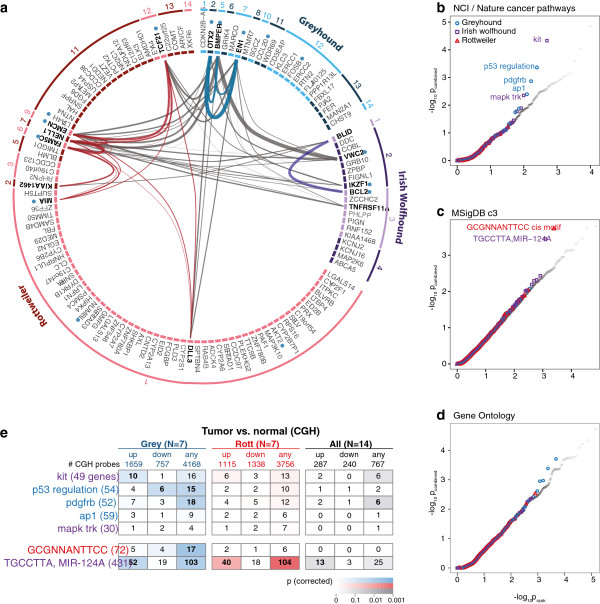
Results: Through genome-wide association analyses in three breeds (greyhounds, Rottweilers, and Irish wolfhounds), we identify 33 inherited risk loci explaining 55% to 85% of phenotype variance in each breed. The greyhound locus exhibiting the strongest association, located 150 kilobases upstream of the genes CDKN2A/B, is also the most rearranged locus in canine osteosarcoma tumors. The top germline candidate variant is found at a >90% frequency in Rottweilers and Irish wolfhounds, and alters an evolutionarily constrained element that we show has strong enhancer activity in human osteosarcoma cells. In all three breeds, osteosarcoma-associated loci and regions of reduced heterozygosity are enriched for genes in pathways connected to bone differentiation and growth. Several pathways, including one of genes regulated by miR124, are also enriched for somatic copy-number changes in tumors.
Conclusions: Mapping a complex cancer in multiple dog breeds reveals a polygenic spectrum of germline risk factors pointing to specific pathways as drivers of disease.
Figures
References
-
- Berman SD, Calo E, Landman AS, Danielian PS, Miller ES, West JC, Fonhoue BD, Caron A, Bronson R, Bouxsein ML, Mukherjee S, Lees JA. Metastatic osteosarcoma induced by inactivation of Rb and p53 in the osteoblast lineage. Proc Natl Acad Sci U S A. 2008;14:11851–11856. doi: 10.1073/pnas.0805462105. - DOI - PMC - PubMed
-
- Walkley CR, Qudsi R, Sankaran VG, Perry JA, Gostissa M, Roth SI, Rodda SJ, Snay E, Dunning P, Fahey FH, Alt FW, McMahon AP, Orkin SH. Conditional mouse osteosarcoma, dependent on p53 loss and potentiated by loss of Rb, mimics the human disease. Genes Dev. 2008;14:1662–1676. doi: 10.1101/gad.1656808. - DOI - PMC - PubMed
-
- Savage SA, Mirabello L, Wang Z, Gastier-Foster JM, Gorlick R, Khanna C, Flanagan AM, Tirabosco R, Andrulis IL, Wunder JS, Gokgoz N, Patino-Garcia A, Sierrasesumaga L, Lecanda F, Kurucu N, Ilhan IE, Sari N, Serra M, Hattinger C, Picci P, Spector LG, Barkauskas DA, Marina N, de Toledo SR, Petrilli AS, Amary MF, Halai D, Thomas DM, Douglass C, Meltzer PS. et al. Genome-wide association study identifies two susceptibility loci for osteosarcoma. Nat Genet. 2013;14:799–803. doi: 10.1038/ng.2645. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
