

The glaucoma-associated olfactomedin domain of myocilin forms polymorphic fibrils that are constrained by partial unfolding and peptide sequence
- PMID: 24333014
- PMCID: PMC3946817
- DOI: 10.1016/j.jmb.2013.12.002
The glaucoma-associated olfactomedin domain of myocilin forms polymorphic fibrils that are constrained by partial unfolding and peptide sequence
Abstract
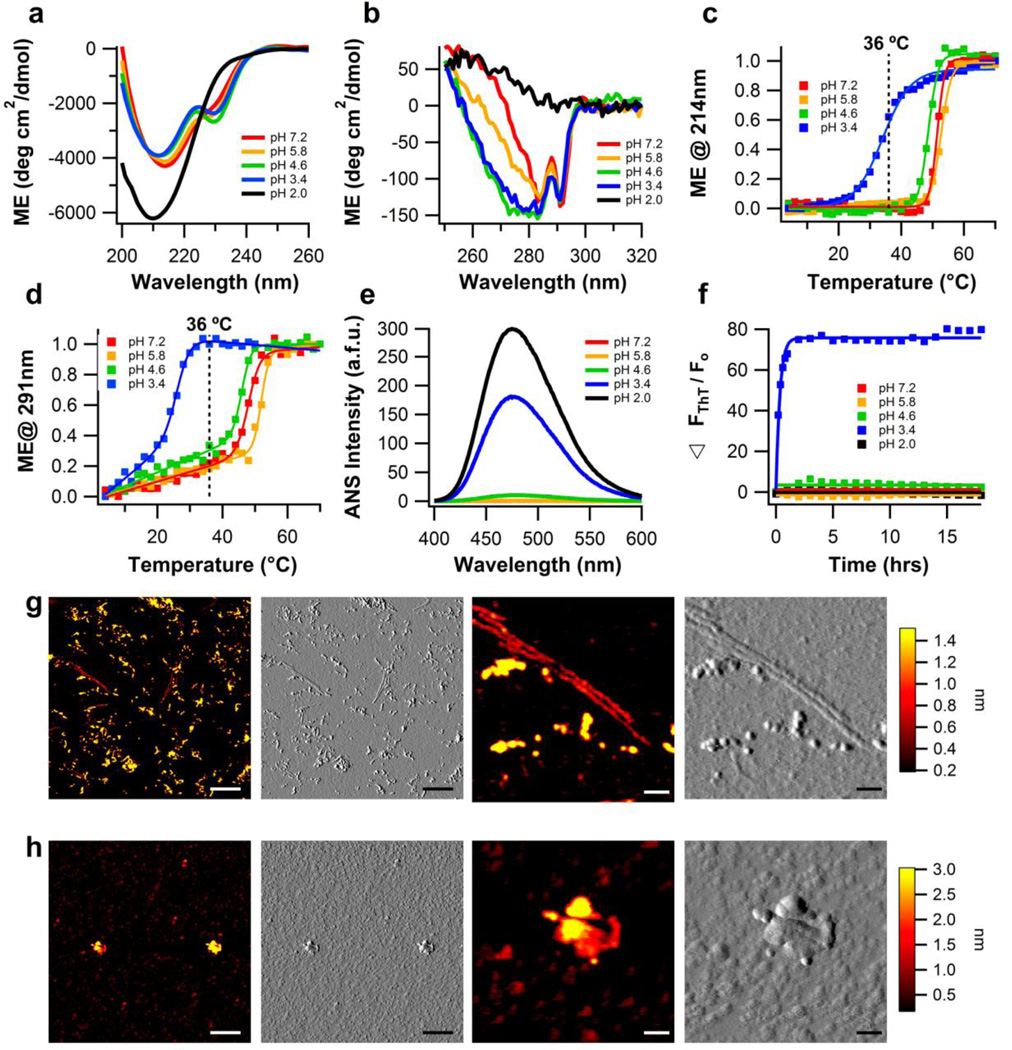
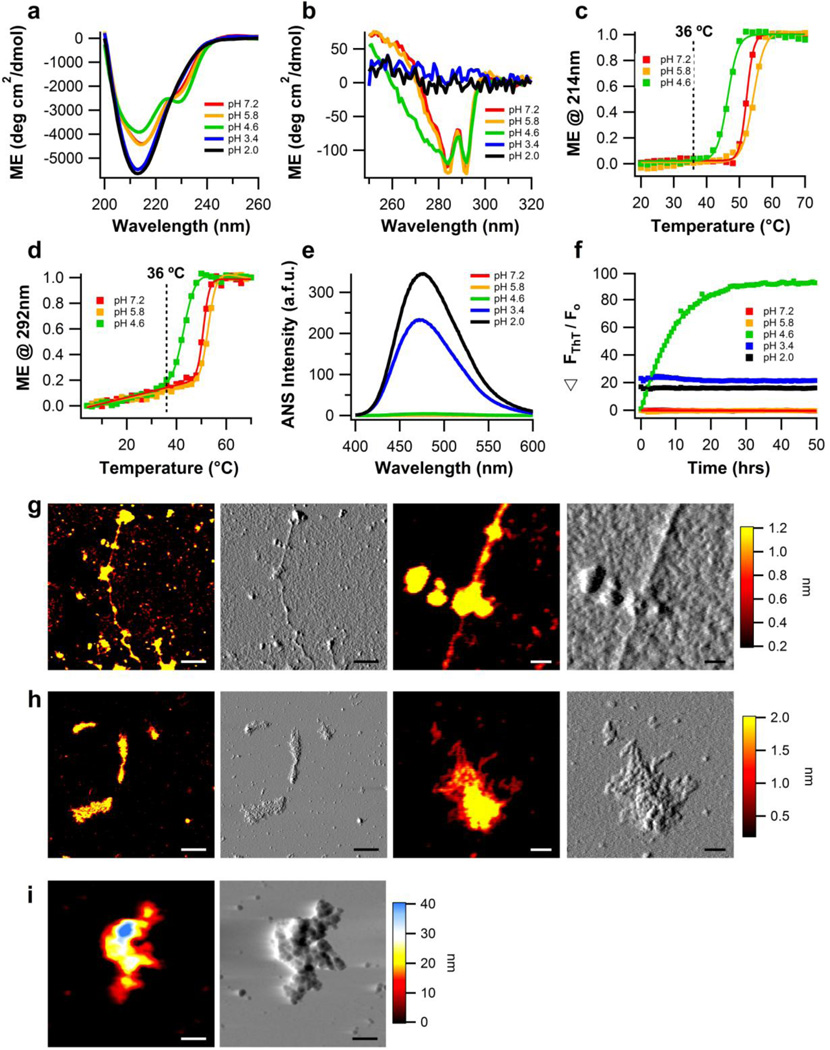
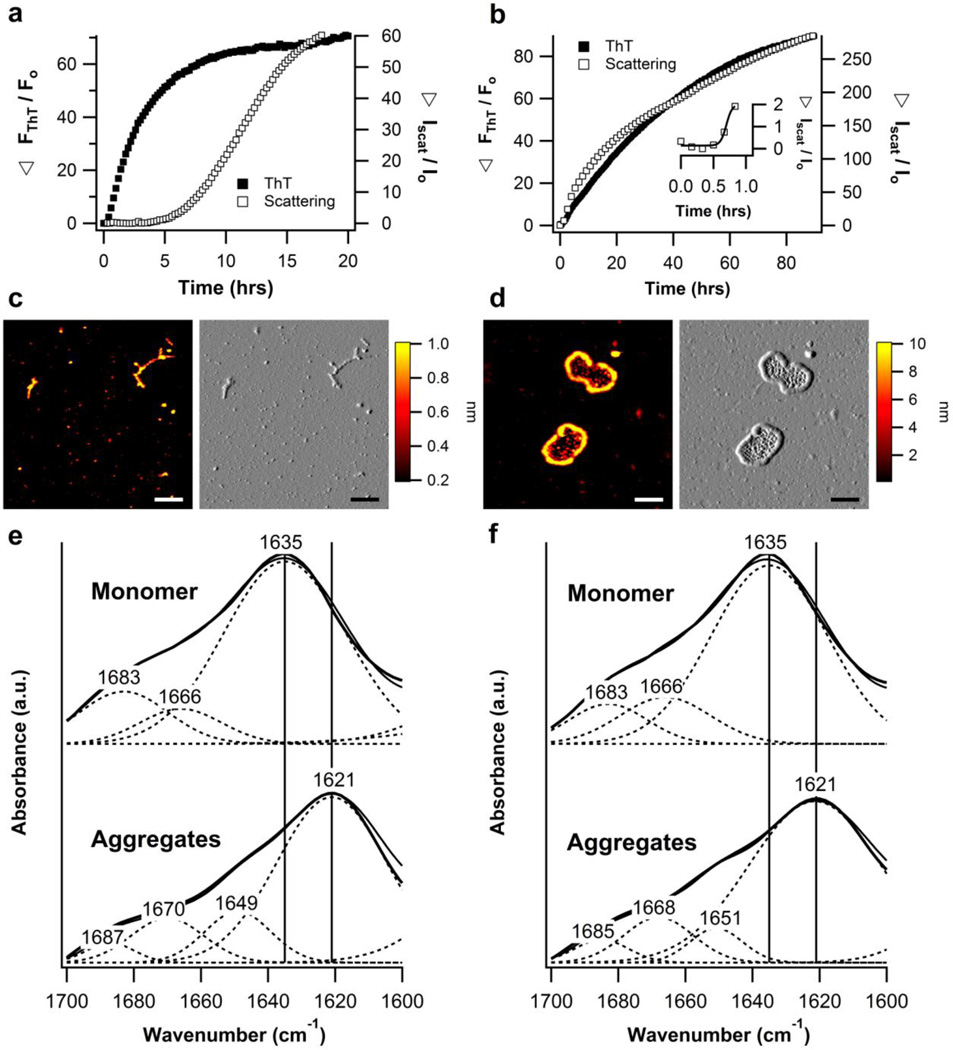
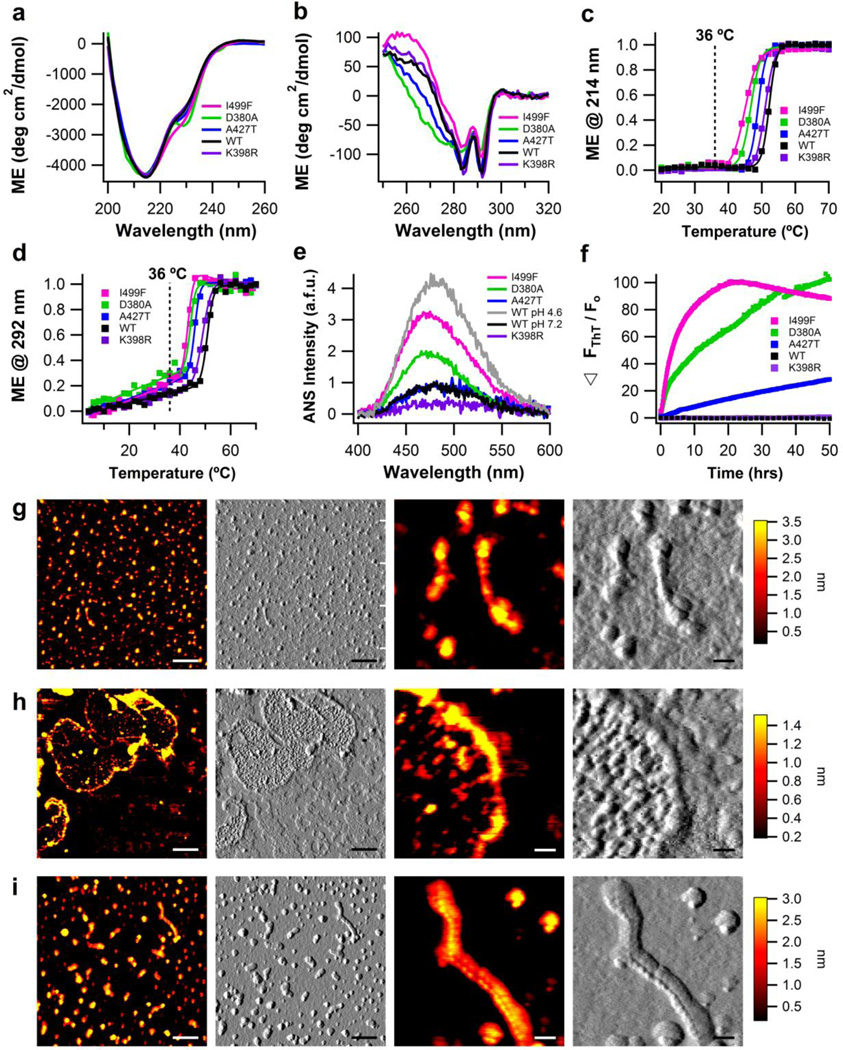
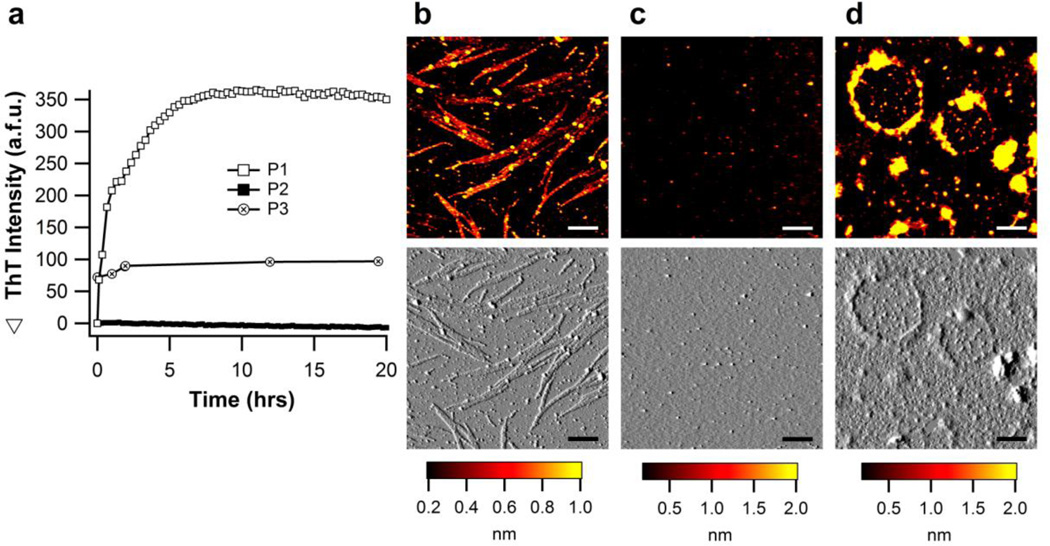
The glaucoma-associated olfactomedin domain of myocilin (myoc-OLF) is a recent addition to the growing list of disease-associated amyloidogenic proteins. Inherited, disease-causing myocilin variants aggregate intracellularly instead of being secreted to the trabecular meshwork, which is a scenario toxic to trabecular meshwork cells and leads to early onset of ocular hypertension, the major risk factor for glaucoma. Here we systematically structurally and biophysically dissected myoc-OLF to better understand its amyloidogenesis. Under mildly destabilizing conditions, wild-type myoc-OLF adopts non-native structures that readily fibrillize when incubated at a temperature just below the transition for tertiary unfolding. With buffers at physiological pH, two main endpoint fibril morphologies are observed: (a) straight fibrils common to many amyloids and (b) unique micron-length, ~300 nm or larger diameter, species that lasso oligomers, which also exhibit classical spectroscopic amyloid signatures. Three disease-causing variants investigated herein exhibit non-native tertiary structures under physiological conditions, leading to a variety of growth rates and a fibril morphologies. In particular, the well-documented D380A variant, which lacks calcium, forms large circular fibrils. Two amyloid-forming peptide stretches have been identified, one for each of the main fibril morphologies observed. Our study places myoc-OLF within the larger landscape of the amylome and provides insight into the diversity of myoc-OLF aggregation that plays a role in glaucoma pathogenesis.
Keywords: AFM; ANS; ER; FTIR; Fourier transform infrared spectroscopy; TEV; TM; ThT; amyloid; anilinonaphthalene-1-sulfonate; atomic force microscopy; circular dichroism; endoplasmic reticulum; protein misfolding; protein structure; thioflavin T; tobacco etch virus; trabecular meshwork.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Stone EM, Fingert JH, Alward WL, Nguyen TD, Polansky JR, Sunden SL, Nishimura D, Clark AF, Nystuen A, Nichols BE, Mackey DA, Ritch R, Kalenak JW, Craven ER, Sheffield VC. Identification of a gene that causes primary open angle glaucoma. Science. 1997;275:668–670. - PubMed
-
- Gobeil S, Rodrigue M-A, Moisan S, Nguyen TD, Polansky JR, Morissette J, Raymond V. Intracellular sequestration of hetero-oligomers formed by wild-type and glaucoma-causing myocilin mutants. Invest. Ophthalmol. Vis. Sci. 2004;45:3560–3567. - PubMed
-
- Joe MK, Sohn S, Hur W, Moon Y, Choi YR, Kee C. Accumulation of mutant myocilins in ER leads to ER stress and potential cytotoxicity in human trabecular meshwork cells. Biochem. Biophys. Res. Commun. 2003;312:592–600. - PubMed
-
- Zhou Z, Vollrath D. A cellular assay distinguishes normal and mutant TIGR/myocilin protein. Hum. Mol. Genet. 1999;8:2221–2218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
