

Trafficking mechanisms of extracellular matrix macromolecules: insights from vertebrate development and human diseases
- PMID: 24333299
- PMCID: PMC3915713
- DOI: 10.1016/j.biocel.2013.11.005
Trafficking mechanisms of extracellular matrix macromolecules: insights from vertebrate development and human diseases
Abstract
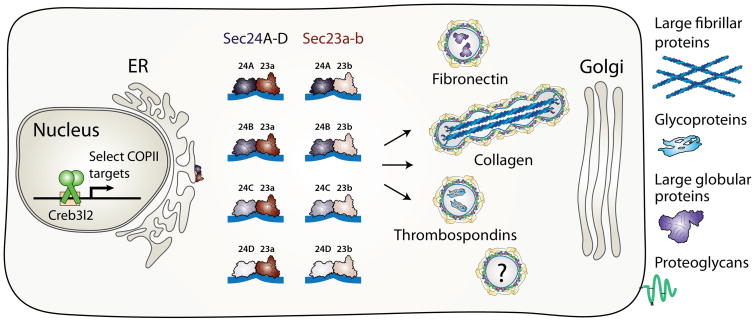
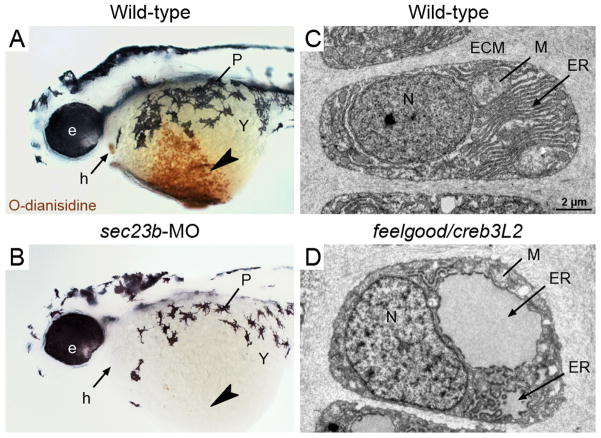
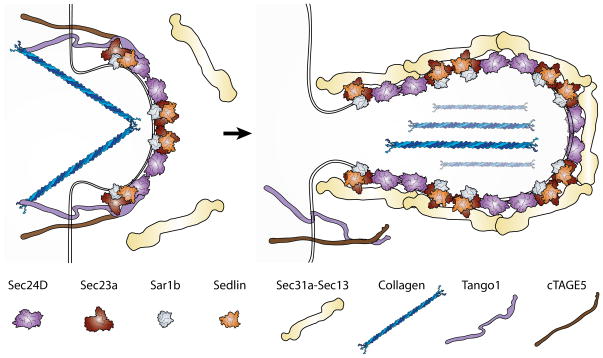
Cellular life depends on protein transport and membrane traffic. In multicellular organisms, membrane traffic is required for extracellular matrix deposition, cell adhesion, growth factor release, and receptor signaling, which are collectively required to integrate the development and physiology of tissues and organs. Understanding the regulatory mechanisms that govern cargo and membrane flow presents a prime challenge in cell biology. Extracellular matrix (ECM) secretion remains poorly understood, although given its essential roles in the regulation of cell migration, differentiation, and survival, ECM secretion mechanisms are likely to be tightly controlled. Recent studies in vertebrate model systems, from fishes to mammals and in human patients, have revealed complex and diverse loss-of-function phenotypes associated with mutations in components of the secretory machinery. A broad spectrum of diseases from skeletal and cardiovascular to neurological deficits have been linked to ECM trafficking. These discoveries have directly challenged the prevailing view of secretion as an essential but monolithic process. Here, we will discuss the latest findings on mechanisms of ECM trafficking in vertebrates.
Keywords: Cartilage and bone; Collagen secretion; ECM; Membrane traffic; Vertebrate animal models.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare that they do not have any competing or financial interests.
Figures
References
-
- Abrams EW, Andrew DJ. CrebA regulates secretory activity in the Drosophila salivary gland and epidermis. Development. 2005;132:2743–58. - PubMed
-
- Angers S, Li T, Yi X, MacCoss MJ, Moon RT, Zheng N. Molecular architecture and assembly of the DDB1-CUL4A ubiquitin ligase machinery. Nature. 2006;443:590–3. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
