

Identification of novel methyltransferases, Bmt5 and Bmt6, responsible for the m3U methylations of 25S rRNA in Saccharomyces cerevisiae
- PMID: 24335083
- PMCID: PMC3950682
- DOI: 10.1093/nar/gkt1281
Identification of novel methyltransferases, Bmt5 and Bmt6, responsible for the m3U methylations of 25S rRNA in Saccharomyces cerevisiae
Abstract
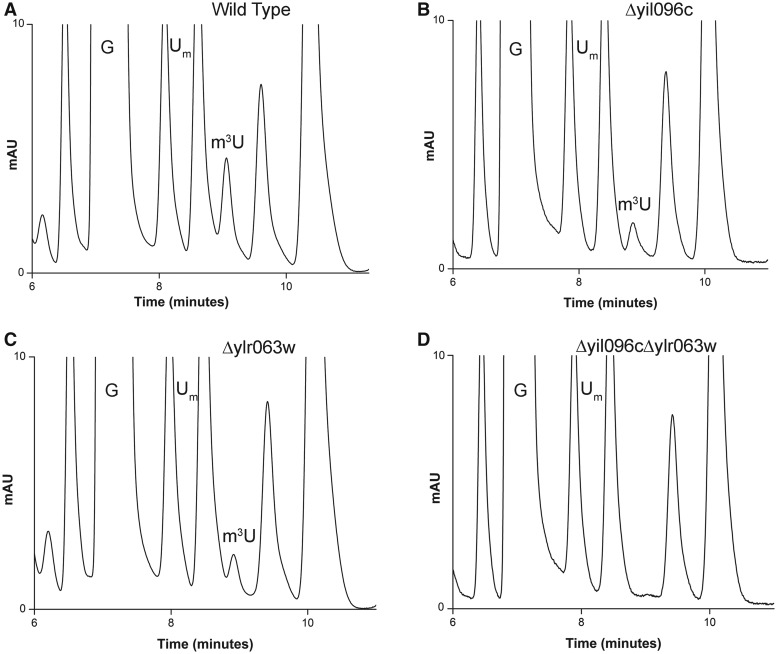
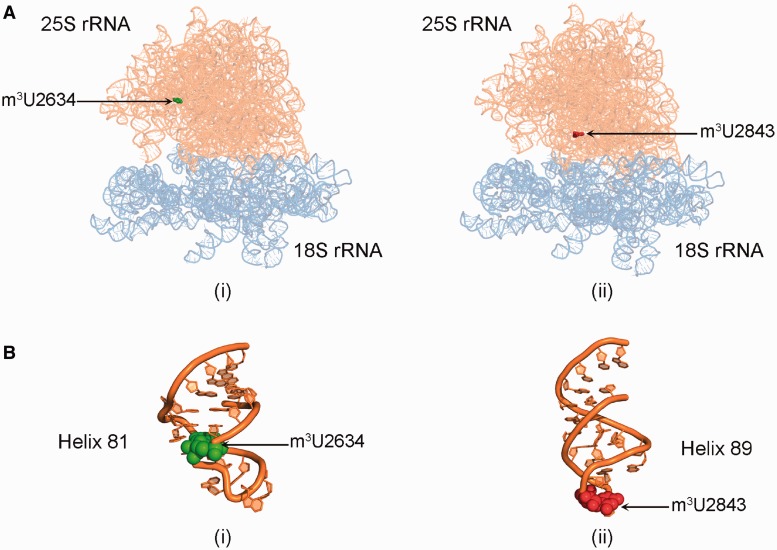
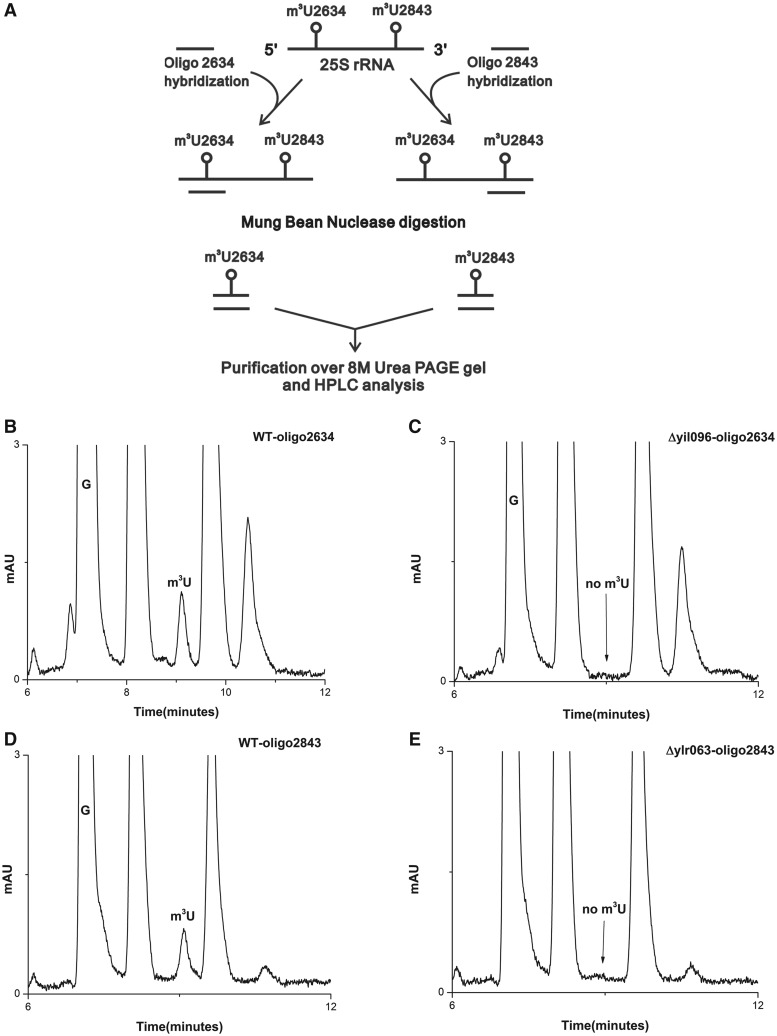
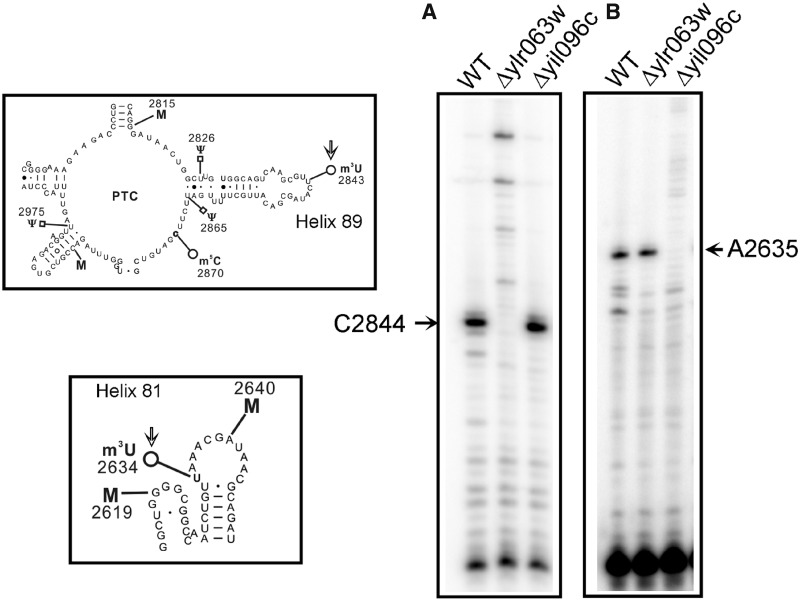
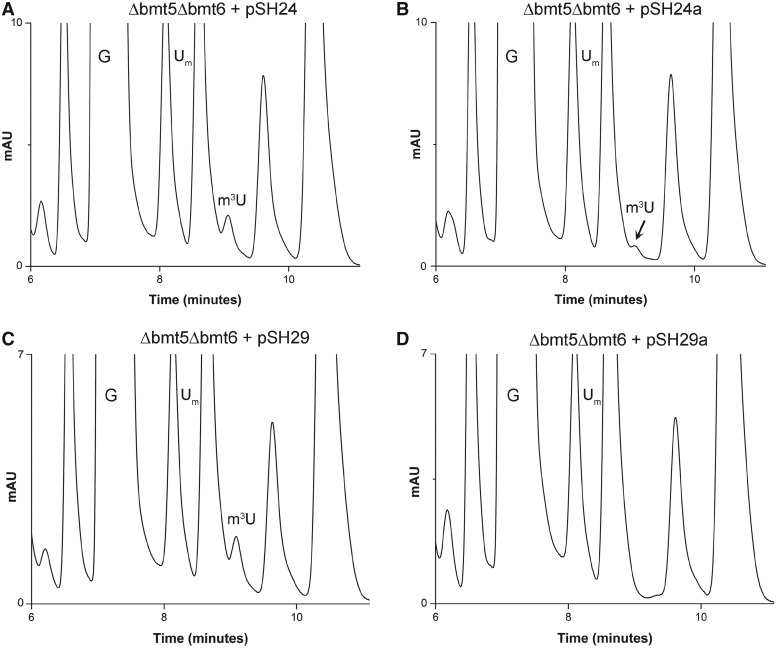
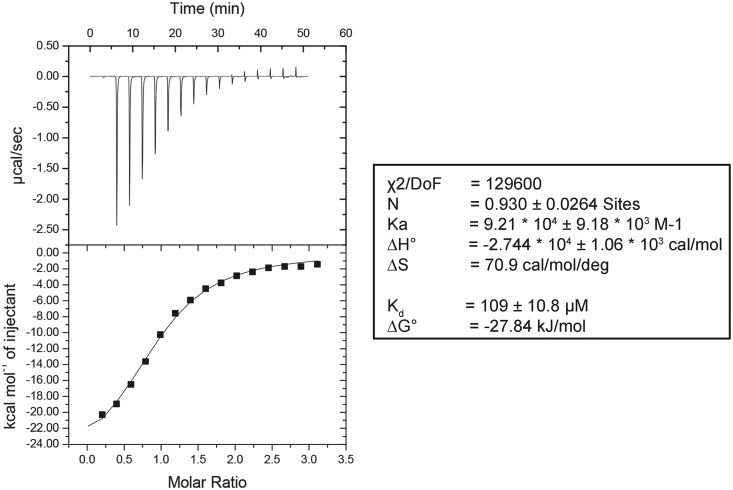
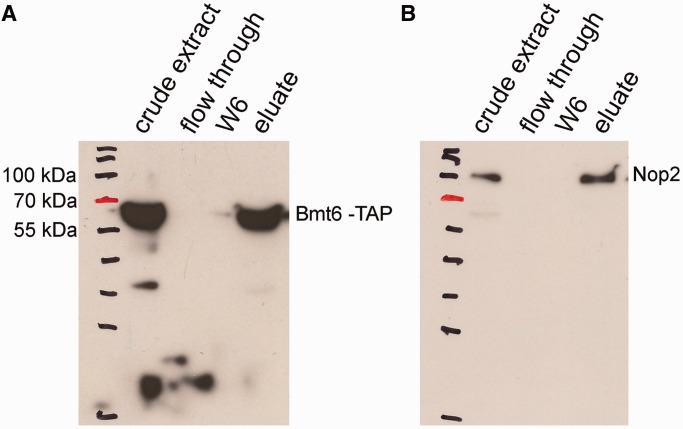
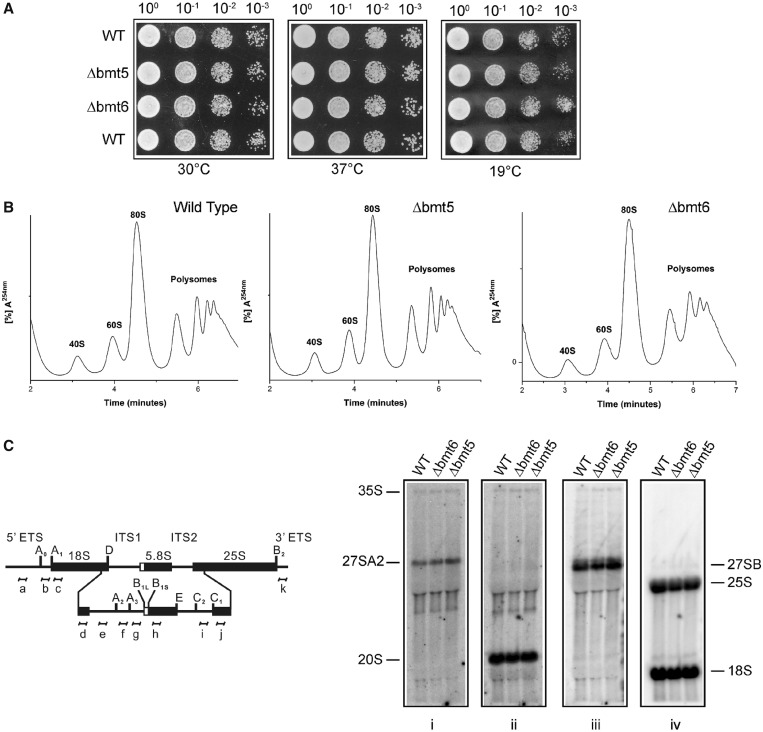
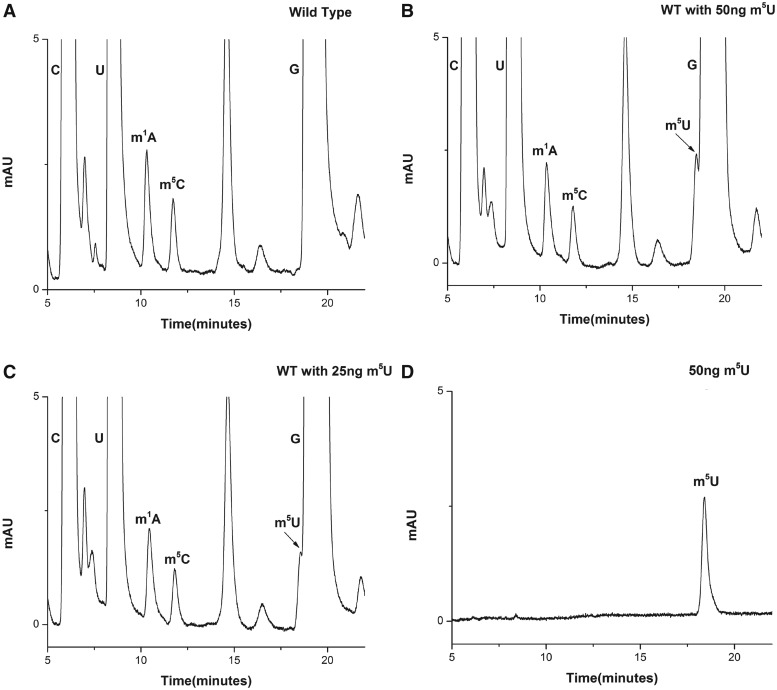
RNA contains various chemical modifications that expand its otherwise limited repertoire to mediate complex processes like translation and gene regulation. 25S rRNA of the large subunit of ribosome contains eight base methylations. Except for the methylation of uridine residues, methyltransferases for all other known base methylations have been recently identified. Here we report the identification of BMT5 (YIL096C) and BMT6 (YLR063W), two previously uncharacterized genes, to be responsible for m3U2634 and m3U2843 methylation of the 25S rRNA, respectively. These genes were identified by RP-HPLC screening of all deletion mutants of putative RNA methyltransferases and were confirmed by gene complementation and phenotypic characterization. Both proteins belong to Rossmann-fold-like methyltransferases and the point mutations in the S-adenosyl-L-methionine binding pocket abolish the methylation reaction. Bmt5 localizes in the nucleolus, whereas Bmt6 is localized predominantly in the cytoplasm. Furthermore, we showed that 25S rRNA of yeast does not contain any m5U residues as previously predicted. With Bmt5 and Bmt6, all base methyltransferases of the 25S rRNA have been identified. This will facilitate the analyses of the significance of these modifications in ribosome function and cellular physiology.
Figures
References
-
- Ben-Shem A, Garreau de Loubresse N, Melnikov S, Jenner L, Yusupova G, Yusupov M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science. 2011;334:1524–1529. - PubMed
-
- Lilley DM. The ribosome functions as a ribozyme. Chembiochem. 2001;2:31–35. - PubMed
-
- Klootwijk J, Planta RJ. Analysis of the methylation sites in yeast ribosomal RNA. Eur. J. Biochem. 1973;39:325–333. - PubMed
-
- Brand RC, Klootwijk J, Van Steenbergen TJ, De Kok AJ, Planta RJ. Secondary methylation of yeast ribosomal precursor RNA. Eur. J. Biochem. 1977;75:311–318. - PubMed
-
- Decatur WA, Fournier MJ. rRNA modifications and ribosome function. Trends Biochem. Sci. 2002;27:344–351. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
