

Rare coding variants in the phospholipase D3 gene confer risk for Alzheimer's disease
- PMID: 24336208
- PMCID: PMC4050701
- DOI: 10.1038/nature12825
Rare coding variants in the phospholipase D3 gene confer risk for Alzheimer's disease
Abstract
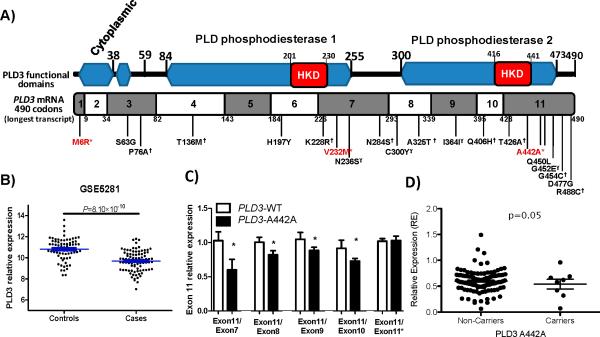
Genome-wide association studies (GWAS) have identified several risk variants for late-onset Alzheimer's disease (LOAD). These common variants have replicable but small effects on LOAD risk and generally do not have obvious functional effects. Low-frequency coding variants, not detected by GWAS, are predicted to include functional variants with larger effects on risk. To identify low-frequency coding variants with large effects on LOAD risk, we carried out whole-exome sequencing (WES) in 14 large LOAD families and follow-up analyses of the candidate variants in several large LOAD case-control data sets. A rare variant in PLD3 (phospholipase D3; Val232Met) segregated with disease status in two independent families and doubled risk for Alzheimer's disease in seven independent case-control series with a total of more than 11,000 cases and controls of European descent. Gene-based burden analyses in 4,387 cases and controls of European descent and 302 African American cases and controls, with complete sequence data for PLD3, reveal that several variants in this gene increase risk for Alzheimer's disease in both populations. PLD3 is highly expressed in brain regions that are vulnerable to Alzheimer's disease pathology, including hippocampus and cortex, and is expressed at significantly lower levels in neurons from Alzheimer's disease brains compared to control brains. Overexpression of PLD3 leads to a significant decrease in intracellular amyloid-β precursor protein (APP) and extracellular Aβ42 and Aβ40 (the 42- and 40-residue isoforms of the amyloid-β peptide), and knockdown of PLD3 leads to a significant increase in extracellular Aβ42 and Aβ40. Together, our genetic and functional data indicate that carriers of PLD3 coding variants have a twofold increased risk for LOAD and that PLD3 influences APP processing. This study provides an example of how densely affected families may help to identify rare variants with large effects on risk for disease or other complex traits.
Figures






Comment in
-
PLD3 and sporadic Alzheimer's disease risk.Nature. 2015 Apr 2;520(7545):E1. doi: 10.1038/nature14036. Nature. 2015. PMID: 25832408 No abstract available.
-
Cruchaga & Goate reply.Nature. 2015 Apr 2;520(7545):E10. doi: 10.1038/nature14041. Nature. 2015. PMID: 25832409 No abstract available.
-
PLD3 variants in population studies.Nature. 2015 Apr 2;520(7545):E2-3. doi: 10.1038/nature14038. Nature. 2015. PMID: 25832410 Free PMC article. No abstract available.
-
PLD3 in non-familial Alzheimer's disease.Nature. 2015 Apr 2;520(7545):E3-5. doi: 10.1038/nature14039. Nature. 2015. PMID: 25832411 No abstract available.
-
Cruchaga & Goate reply.Nature. 2015 Apr 2;520(7545):E5-6. doi: 10.1038/nature14037. Nature. 2015. PMID: 25832412 No abstract available.
-
PLD3 gene variants and Alzheimer's disease.Nature. 2015 Apr 2;520(7545):E7-8. doi: 10.1038/nature14040. Nature. 2015. PMID: 25832413 No abstract available.
References
-
- Bertram L, McQueen M, Mullin K, Blacker D, Tanzi R. [1/26/2013];The AlzGene Database. Alzheimer Research Forum. Available at: http://www.alzgene.org.
-
- Goate A, Hardy J. Twenty years of Alzheimer's disease-causing mutations. J Neurochem. 2012;120(Suppl 1):3–8. doi:10.1111/j.1471-4159.2011.07575.x. - PubMed
-
- Jonsson T, et al. A mutation in APP protects against Alzheimer's disease and age-related cognitive decline. Nature. 2012;488:96–99. doi:10.1038/nature11283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- G0802189/MRC_/Medical Research Council/United Kingdom
- R01 AG042611/AG/NIA NIH HHS/United States
- ZO1 AG000950-10/AG/NIA NIH HHS/United States
- CAPMC/ CIHR/Canada
- U24 AG026395/AG/NIA NIH HHS/United States
- R01 AG041797/AG/NIA NIH HHS/United States
- R01 AG030653/AG/NIA NIH HHS/United States
- R01 AG021136/AG/NIA NIH HHS/United States
- 100140/WT_/Wellcome Trust/United Kingdom
- R01 AG039700/AG/NIA NIH HHS/United States
- G0901254/MRC_/Medical Research Council/United Kingdom
- ZO1AG000950-11/AG/NIA NIH HHS/United States
- R01AG21136/AG/NIA NIH HHS/United States
- R01-AG042611/AG/NIA NIH HHS/United States
- R01-AG11380/AG/NIA NIH HHS/United States
- AG07562/AG/NIA NIH HHS/United States
- 1R01AG041797/AG/NIA NIH HHS/United States
- R01 AG044546/AG/NIA NIH HHS/United States
- U24 AG021886/AG/NIA NIH HHS/United States
- MR/L016400/1/MRC_/Medical Research Council/United Kingdom
- P01 AG003991/AG/NIA NIH HHS/United States
- P50 AG005681/AG/NIA NIH HHS/United States
- U24 AG056270/AG/NIA NIH HHS/United States
- R01-AG21136/AG/NIA NIH HHS/United States
- 5U24AG026395/AG/NIA NIH HHS/United States
- P01 AG026276/AG/NIA NIH HHS/United States
- P30-NS069329/NS/NINDS NIH HHS/United States
- U24AG21886/AG/NIA NIH HHS/United States
- P01 AG03991/AG/NIA NIH HHS/United States
- MC_G1000735/MRC_/Medical Research Council/United Kingdom
- NIH R01039700/PHS HHS/United States
- Z01 AG000950/ImNIH/Intramural NIH HHS/United States
- 089703/WT_/Wellcome Trust/United Kingdom
- AG041718/AG/NIA NIH HHS/United States
- G1100695/MRC_/Medical Research Council/United Kingdom
- NIH P50 AG05681/AG/NIA NIH HHS/United States
- 089698/WT_/Wellcome Trust/United Kingdom
- R01-AG18712/AG/NIA NIH HHS/United States
- K01 AG046374/AG/NIA NIH HHS/United States
- AG023652/AG/NIA NIH HHS/United States
- R01-AG035083/AG/NIA NIH HHS/United States
- AG030653/AG/NIA NIH HHS/United States
- R01-AG044546/AG/NIA NIH HHS/United States
- P50 AG005133/AG/NIA NIH HHS/United States
- ZIA AG000950/ImNIH/Intramural NIH HHS/United States
- RF1 AG044546/AG/NIA NIH HHS/United States
- R01 AG041718/AG/NIA NIH HHS/United States
- R25 DA027995/DA/NIDA NIH HHS/United States
- P30 NS069329/NS/NINDS NIH HHS/United States
- AG005133/AG/NIA NIH HHS/United States
- 081864/WT_/Wellcome Trust/United Kingdom
- R01 AG011380/AG/NIA NIH HHS/United States
- MC_G1000734/MRC_/Medical Research Council/United Kingdom
- R01 AG035083/AG/NIA NIH HHS/United States
- R01 AG018712/AG/NIA NIH HHS/United States
- G0802462/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
