

Sterilization of granulomas is common in active and latent tuberculosis despite within-host variability in bacterial killing
- PMID: 24336248
- PMCID: PMC3947310
- DOI: 10.1038/nm.3412
Sterilization of granulomas is common in active and latent tuberculosis despite within-host variability in bacterial killing
Abstract
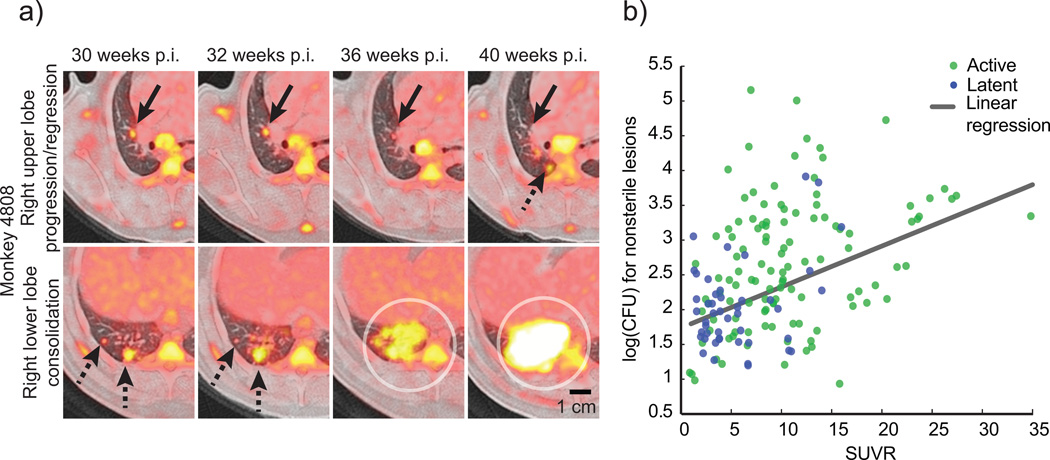
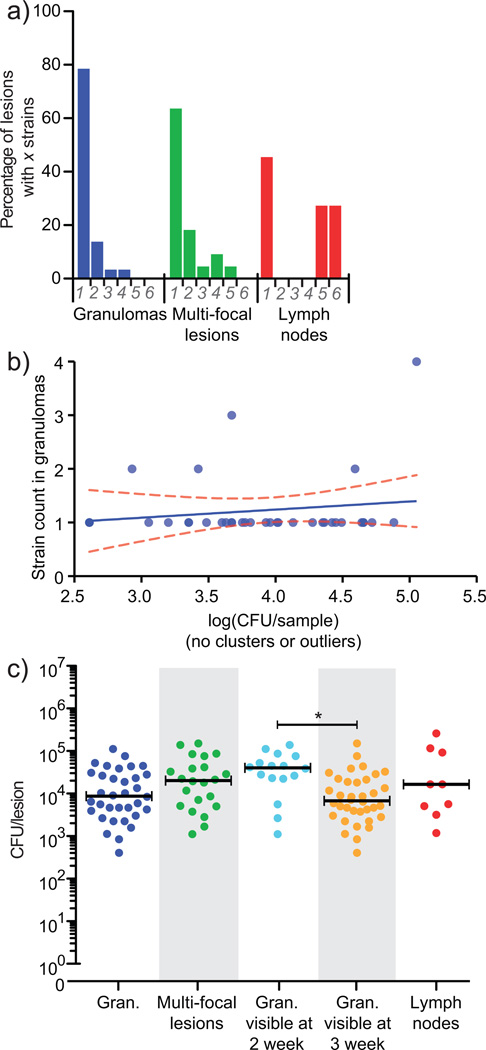
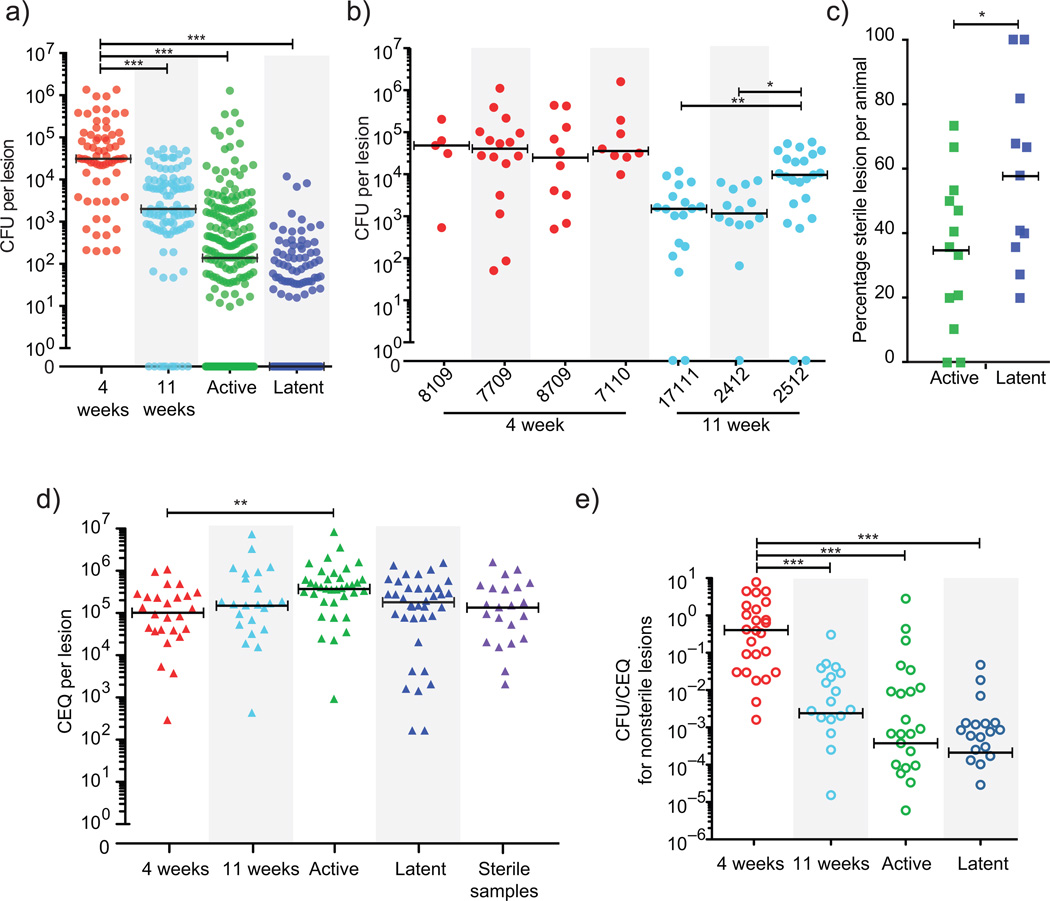
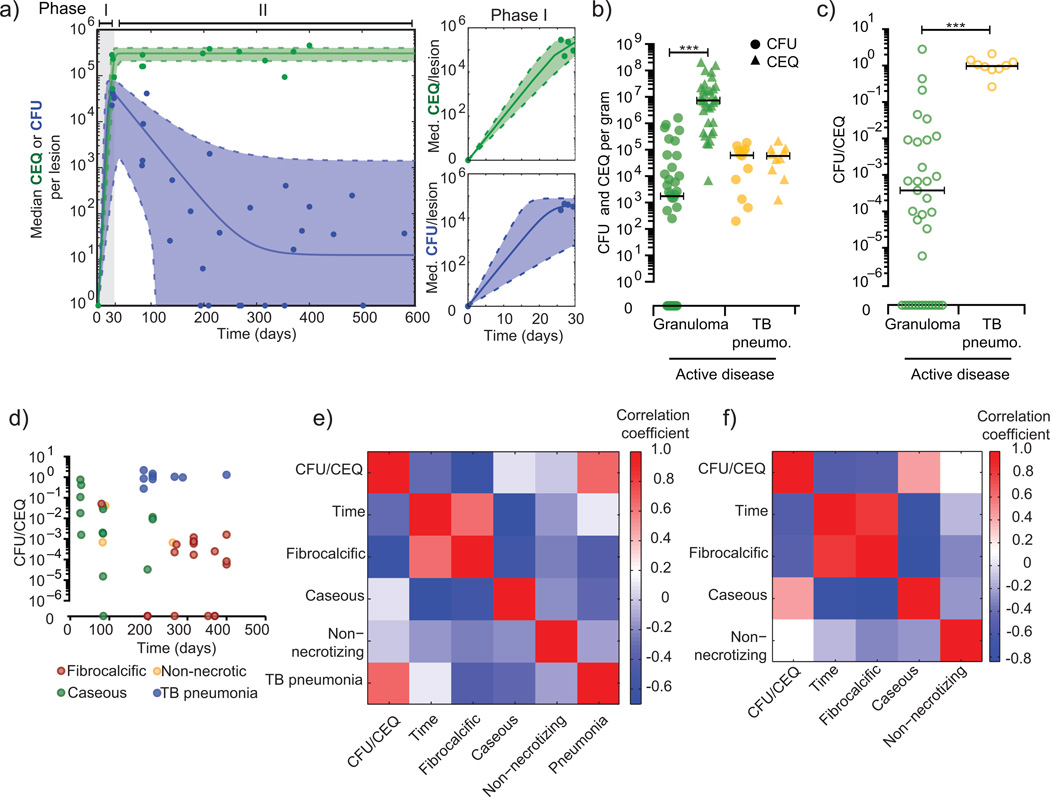
Over 30% of the world's population is infected with Mycobacterium tuberculosis (Mtb), yet only ∼5-10% will develop clinical disease. Despite considerable effort, researchers understand little about what distinguishes individuals whose infection progresses to active tuberculosis (TB) from those whose infection remains latent for decades. The variable course of disease is recapitulated in cynomolgus macaques infected with Mtb. Active disease occurs in ∼45% of infected macaques and is defined by clinical, microbiologic and immunologic signs, whereas the remaining infected animals are clinically asymptomatic. Here, we use individually marked Mtb isolates and quantitative measures of culturable and cumulative bacterial burden to show that most lung lesions are probably founded by a single bacterium and reach similar maximum burdens. Despite this observation, the fate of individual lesions varies substantially within the same host. Notably, in active disease, the host sterilizes some lesions even while others progress. Our data suggest that lesional heterogeneity arises, in part, through differential killing of bacteria after the onset of adaptive immunity. Thus, individual lesions follow diverse and overlapping trajectories, suggesting that critical responses occur at a lesional level to ultimately determine the clinical outcome of infection. Defining the local factors that dictate outcome will be useful in developing effective interventions to prevent active TB.
Figures
References
-
- Zumla A, Raviglione M, Hafner R, von Reyn CF. Tuberculosis. N Engl J Med. 2013;368:745–755. - PubMed
Online References
-
- Kennedy HE, Vandiviere HM, Willis HS. The effects of extended incubation on propagability of tubercle bacilli. Am Rev Tuberc. 1958;77:802–814. - PubMed
-
- Vandiviere HM, Loring WE, Melvin I, Willis S. The treated pulmonary lesion and its tubercle bacillus. II. The death and resurrection. Am J Med Sci. 1956;232:30–37. passim. - PubMed
-
- Keane J, et al. Tuberculosis associated with infliximab, a tumor necrosis factor alpha-neutralizing agent. N Engl J Med. 2001;345:1098–1104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
