

Genome-wide localization of small molecules
- PMID: 24336317
- PMCID: PMC4189815
- DOI: 10.1038/nbt.2776
Genome-wide localization of small molecules
Abstract
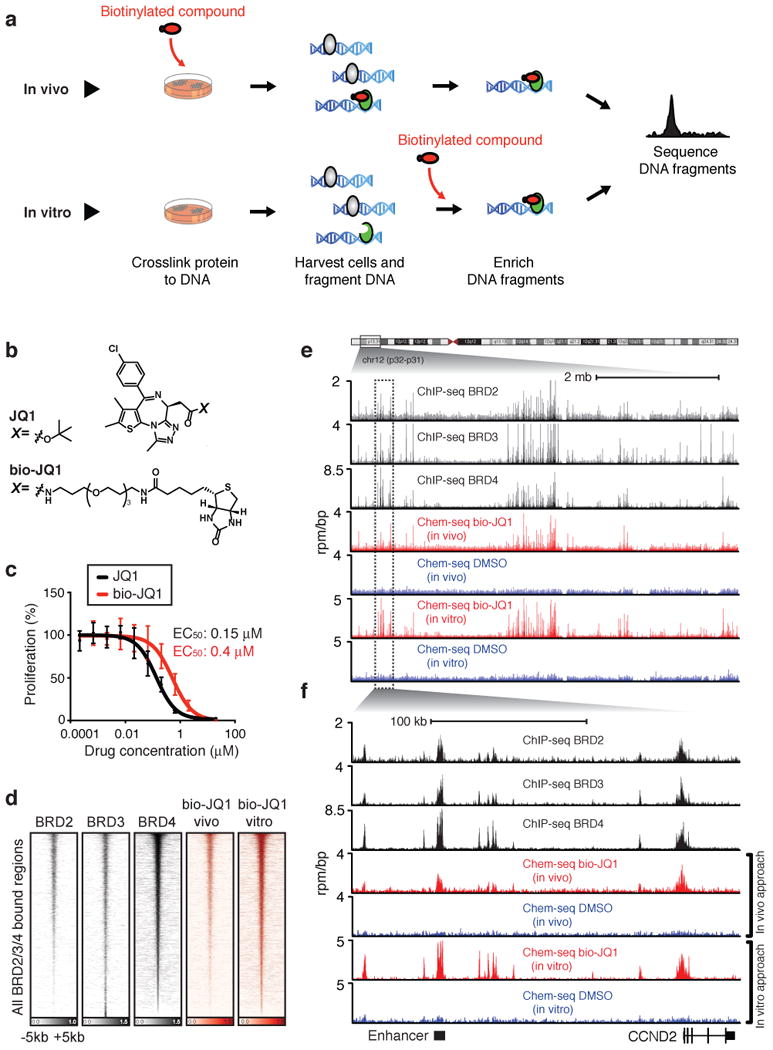
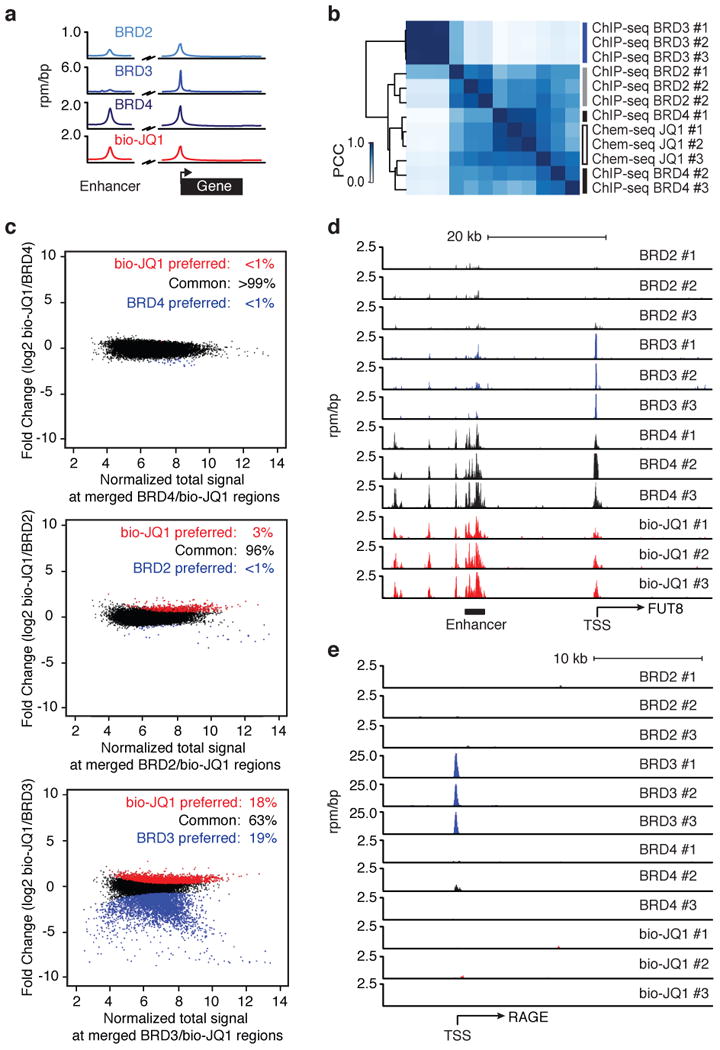
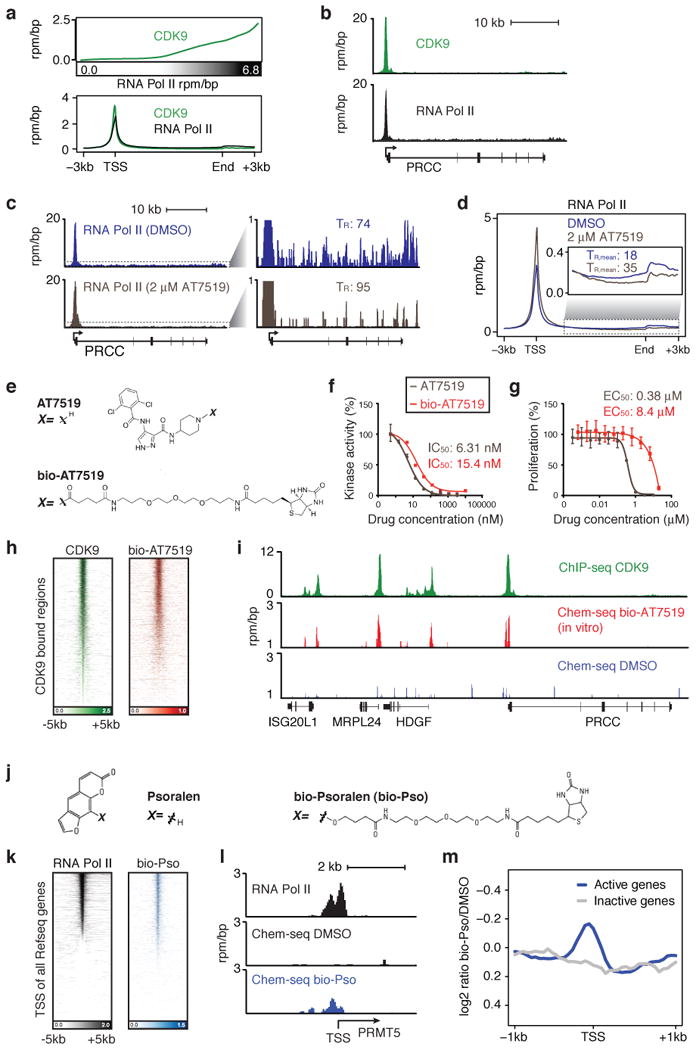
A vast number of small-molecule ligands, including therapeutic drugs under development and in clinical use, elicit their effects by binding specific proteins associated with the genome. An ability to map the direct interactions of a chemical entity with chromatin genome-wide could provide important insights into chemical perturbation of cellular function. Here we describe a method that couples ligand-affinity capture and massively parallel DNA sequencing (Chem-seq) to identify the sites bound by small chemical molecules throughout the human genome. We show how Chem-seq can be combined with ChIP-seq to gain unique insights into the interaction of drugs with their target proteins throughout the genome of tumor cells. These methods will be broadly useful to enhance understanding of therapeutic action and to characterize the specificity of chemical entities that interact with DNA or genome-associated proteins.
Figures
References
-
- Bell O, Tiwari VK, Thoma NH, Schubeler D. Determinants and dynamics of genome accessibility. Nat Rev Genet. 2011;12:554–564. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
