

The Coffin-Lowry syndrome-associated protein RSK2 regulates neurite outgrowth through phosphorylation of phospholipase D1 (PLD1) and synthesis of phosphatidic acid
- PMID: 24336713
- PMCID: PMC6618760
- DOI: 10.1523/JNEUROSCI.2283-13.2013
The Coffin-Lowry syndrome-associated protein RSK2 regulates neurite outgrowth through phosphorylation of phospholipase D1 (PLD1) and synthesis of phosphatidic acid
Abstract
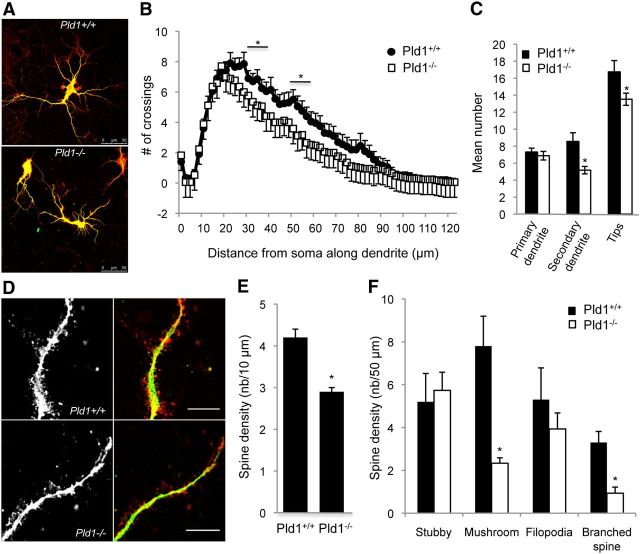
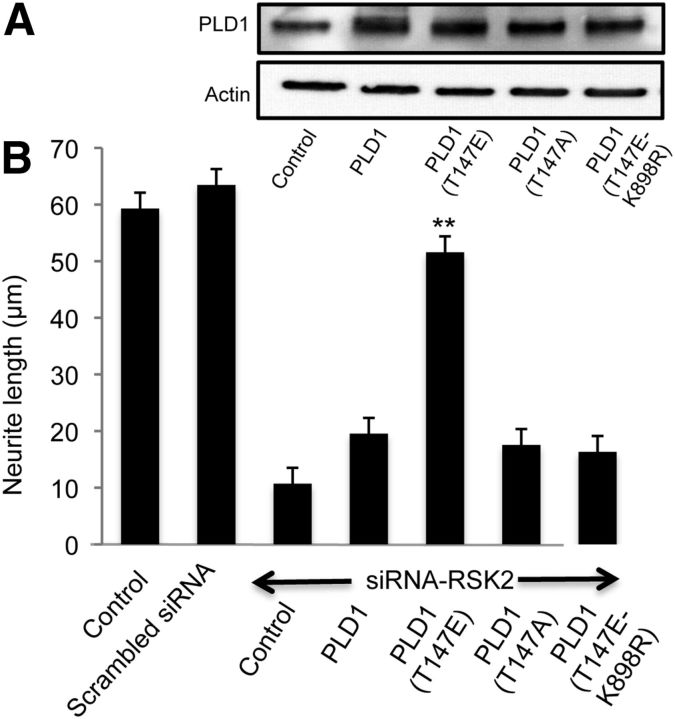
More than 80 human X-linked genes have been associated with mental retardation and deficits in learning and memory. However, most of the identified mutations induce limited morphological alterations in brain organization and the molecular bases underlying neuronal clinical features remain elusive. We show here that neurons cultured from mice lacking ribosomal S6 kinase 2 (Rsk2), a model for the Coffin-Lowry syndrome (CLS), exhibit a significant delay in growth in a similar way to that shown by neurons cultured from phospholipase D1 (Pld1) knock-out mice. We found that gene silencing of Pld1 or Rsk2 as well as acute pharmacological inhibition of PLD1 or RSK2 in PC12 cells strongly impaired neuronal growth factor (NGF)-induced neurite outgrowth. Expression of a phosphomimetic PLD1 mutant rescued the inhibition of neurite outgrowth in PC12 cells silenced for RSK2, revealing that PLD1 is a major target for RSK2 in neurite formation. NGF-triggered RSK2-dependent phosphorylation of PLD1 led to its activation and the synthesis of phosphatidic acid at sites of neurite growth. Additionally, total internal reflection fluorescence microscopy experiments revealed that RSK2 and PLD1 positively control fusion of tetanus neurotoxin insensitive vesicle-associated membrane protein (TiVAMP)/VAMP-7 vesicles at sites of neurite outgrowth. We propose that the loss of function mutations in RSK2 that leads to CLS and neuronal deficits are related to defects in neuronal growth due to impaired RSK2-dependent PLD1 activity resulting in a reduced vesicle fusion rate and membrane supply.
Figures







References
-
- Cai D, Zhong M, Wang R, Netzer WJ, Shields D, Zheng H, Sisodia SS, Foster DA, Gorelick FS, Xu H, Greengard P. Phospholipase D1 corrects impaired betaAPP trafficking and neurite outgrowth in familial Alzheimer's disease-linked presenilin-1 mutant neurons. Proc Natl Acad Sci U S A. 2006;103:1936–1940. doi: 10.1073/pnas.0510710103. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases