

Two small (p)ppGpp synthases in Staphylococcus aureus mediate tolerance against cell envelope stress conditions
- PMID: 24336937
- PMCID: PMC3911181
- DOI: 10.1128/JB.01201-13
Two small (p)ppGpp synthases in Staphylococcus aureus mediate tolerance against cell envelope stress conditions
Abstract
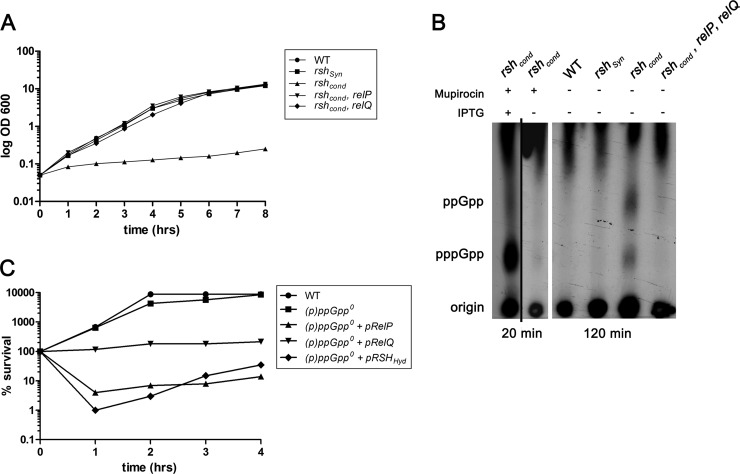
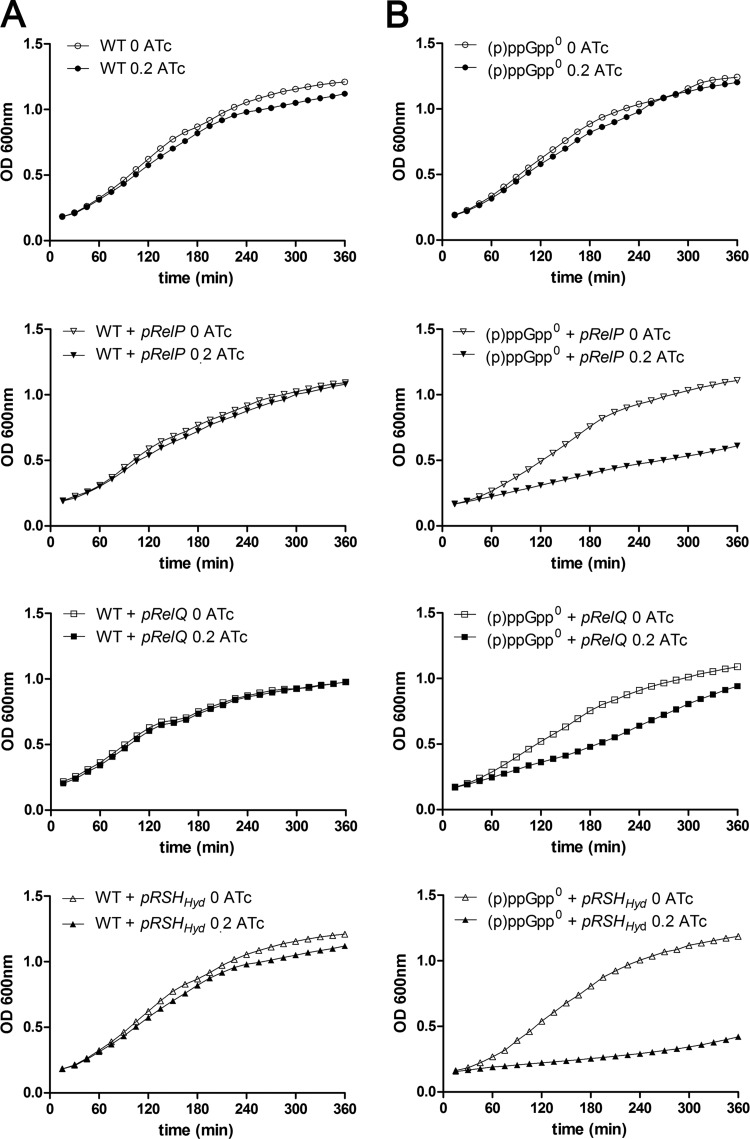
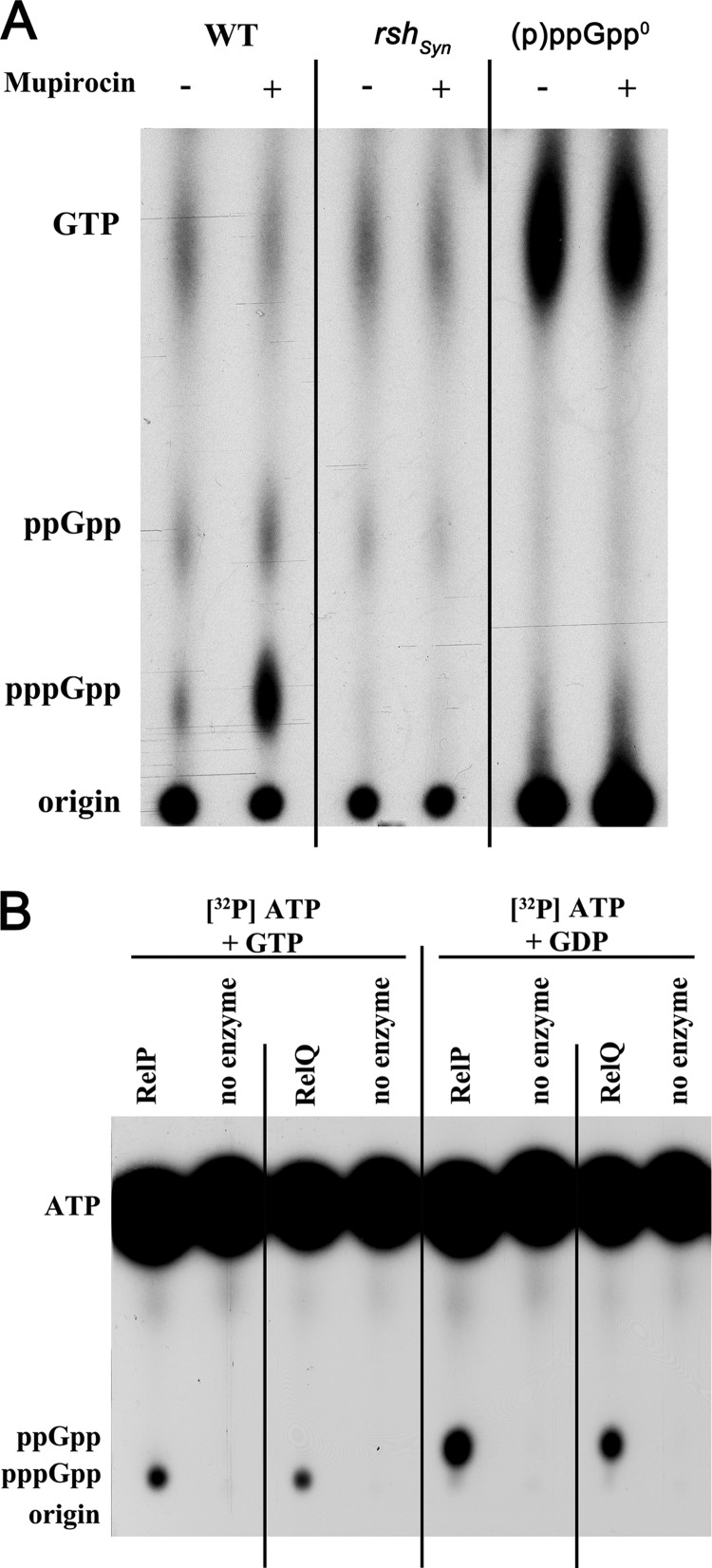
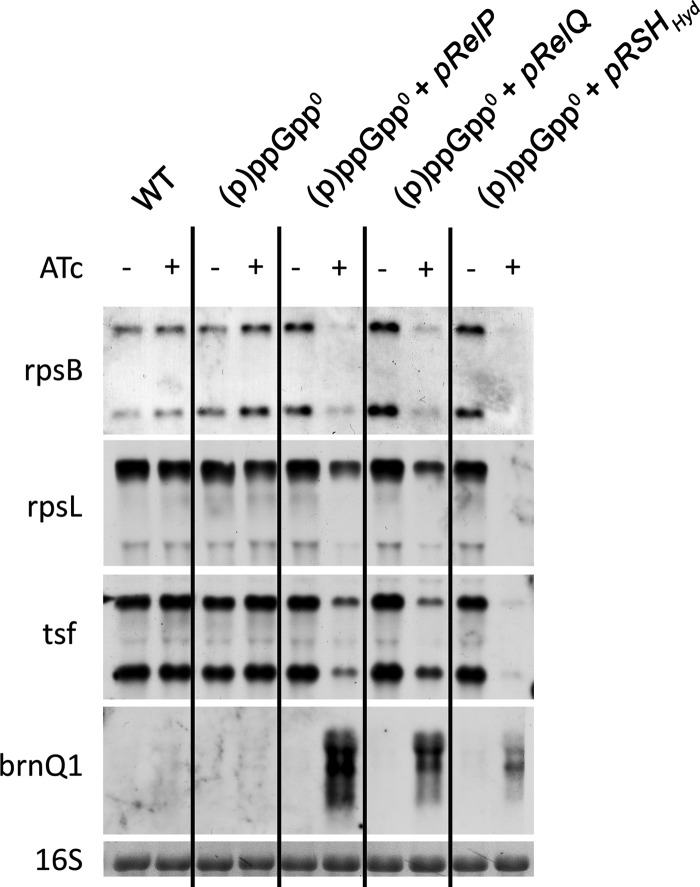
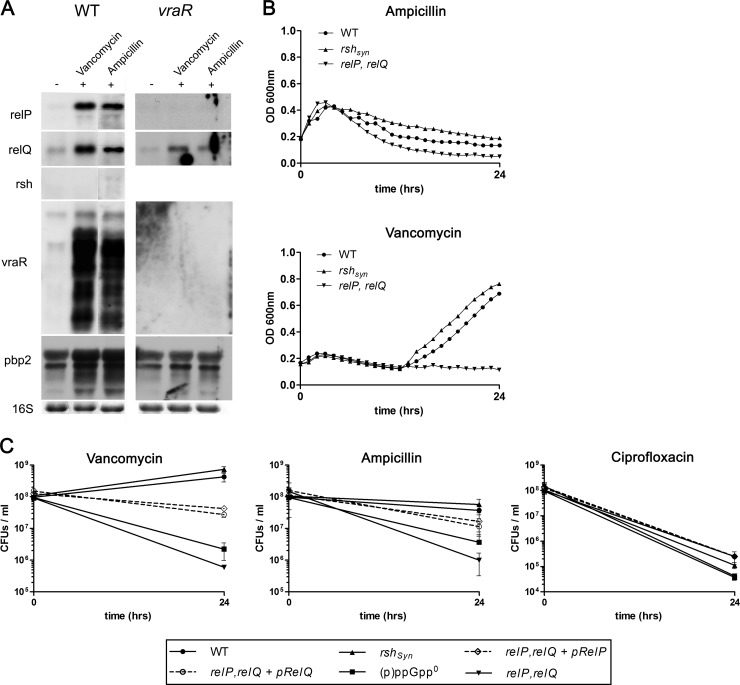
The stringent response is a conserved global regulatory mechanism that is related to the synthesis of (p)ppGpp nucleotides. Gram-positive bacteria, such as Staphylococcus aureus, possess three (p)ppGpp synthases: the bifunctional RSH (RelA/SpoT homolog) protein, which consists of a (p)ppGpp synthase and a (p)ppGpp hydrolase domain, and two truncated (p)ppGpp synthases, designated RelP and RelQ. Here, we characterized these two small (p)ppGpp synthases. Biochemical analyses of purified proteins and in vivo studies revealed a stronger synthetic activity for RelP than for RelQ. However, both enzymes prefer GDP over GTP as the pyrophosphate recipient to synthesize ppGpp. Each of the enzymes was shown to be responsible for the essentiality of the (p)ppGpp hydrolase domain of the RSH protein. The staphylococcal RSH-hydrolase is an efficient enzyme that prevents the toxic accumulation of (p)ppGpp. Expression of (p)ppGpp synthases in a hydrolase-negative background leads not only to growth arrest but also to cell death. Transcriptional analyses showed that relP and relQ are strongly induced upon vancomycin and ampicillin treatments. Accordingly, mutants lacking relP and relQ showed a significantly reduced survival rate upon treatments with cell wall-active antibiotics. Thus, RelP and RelQ are active (p)ppGpp synthases in S. aureus that are induced under cell envelope stress to mediate tolerance against these conditions.
Figures
Similar articles
-
The Ps and Qs of alarmone synthesis in Staphylococcus aureus.PLoS One. 2019 Oct 15;14(10):e0213630. doi: 10.1371/journal.pone.0213630. eCollection 2019. PLoS One. 2019. PMID: 31613897 Free PMC article.
-
Regulation of the opposing (p)ppGpp synthetase and hydrolase activities in a bifunctional RelA/SpoT homologue from Staphylococcus aureus.PLoS Genet. 2018 Jul 9;14(7):e1007514. doi: 10.1371/journal.pgen.1007514. eCollection 2018 Jul. PLoS Genet. 2018. PMID: 29985927 Free PMC article.
-
Structural basis for (p)ppGpp synthesis by the Staphylococcus aureus small alarmone synthetase RelP.J Biol Chem. 2018 Mar 2;293(9):3254-3264. doi: 10.1074/jbc.RA117.001374. Epub 2018 Jan 11. J Biol Chem. 2018. PMID: 29326162 Free PMC article.
-
Triggering the stringent response: signals responsible for activating (p)ppGpp synthesis in bacteria.Microbiology (Reading). 2018 Mar;164(3):268-276. doi: 10.1099/mic.0.000621. Microbiology (Reading). 2018. PMID: 29493495 Review.
-
Within and beyond the stringent response-RSH and (p)ppGpp in plants.Planta. 2017 Nov;246(5):817-842. doi: 10.1007/s00425-017-2780-y. Epub 2017 Sep 25. Planta. 2017. PMID: 28948393 Free PMC article. Review.
Cited by
-
RNA Sequencing Identifies a Common Physiology in Vancomycin- and Ciprofloxacin-Tolerant Staphylococcus aureus Induced by ileS Mutations.Antimicrob Agents Chemother. 2020 Sep 21;64(10):e00827-20. doi: 10.1128/AAC.00827-20. Print 2020 Sep 21. Antimicrob Agents Chemother. 2020. PMID: 32690649 Free PMC article.
-
The (p)ppGpp Synthetase RSH Mediates Stationary-Phase Onset and Antibiotic Stress Survival in Clostridioides difficile.J Bacteriol. 2020 Sep 8;202(19):e00377-20. doi: 10.1128/JB.00377-20. Print 2020 Sep 8. J Bacteriol. 2020. PMID: 32661079 Free PMC article.
-
Stress-induced inactivation of the Staphylococcus aureus purine biosynthesis repressor leads to hypervirulence.Nat Commun. 2019 Feb 15;10(1):775. doi: 10.1038/s41467-019-08724-x. Nat Commun. 2019. PMID: 30770821 Free PMC article.
-
Tale of Twin Bifunctional Second Messenger (p)ppGpp Synthetases and Their Function in Mycobacteria.ACS Omega. 2023 Aug 25;8(36):32258-32270. doi: 10.1021/acsomega.3c03557. eCollection 2023 Sep 12. ACS Omega. 2023. PMID: 37720788 Free PMC article. Review.
-
Subinhibitory Concentrations of Bacteriostatic Antibiotics Induce relA-Dependent and relA-Independent Tolerance to β-Lactams.Antimicrob Agents Chemother. 2017 Mar 24;61(4):e02173-16. doi: 10.1128/AAC.02173-16. Print 2017 Apr. Antimicrob Agents Chemother. 2017. PMID: 28115345 Free PMC article.
References
-
- Xiao H, Kalman M, Ikehara K, Zemel S, Glaser G, Cashel M. 1991. Residual guanosine 3′,5′-bispyrophosphate synthetic activity of relA null mutants can be eliminated by spoT null mutations. J. Biol. Chem. 266:5980–5990 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
