

Lysosomal multienzyme complex: pros and cons of working together
- PMID: 24337808
- PMCID: PMC4037752
- DOI: 10.1007/s00018-013-1538-3
Lysosomal multienzyme complex: pros and cons of working together
Abstract
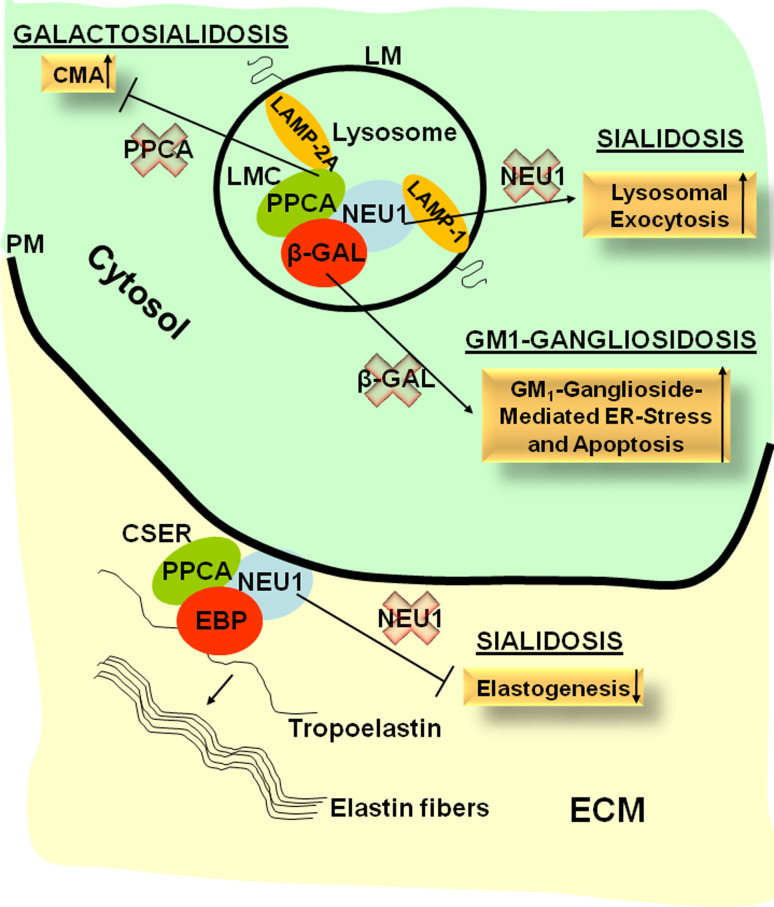
The ubiquitous distribution of lysosomes and their heterogeneous protein composition reflects the versatility of these organelles in maintaining cell homeostasis and their importance in tissue differentiation and remodeling. In lysosomes, the degradation of complex, macromolecular substrates requires the synergistic action of multiple hydrolases that usually work in a stepwise fashion. This catalytic machinery explains the existence of lysosomal enzyme complexes that can be dynamically assembled and disassembled to efficiently and quickly adapt to the pool of substrates to be processed or degraded, adding extra tiers to the regulation of the individual protein components. An example of such a complex is the one composed of three hydrolases that are ubiquitously but differentially expressed: the serine carboxypeptidase, protective protein/cathepsin A (PPCA), the sialidase, neuraminidase-1 (NEU1), and the glycosidase β-galactosidase (β-GAL). Next to this 'core' complex, the existence of sub-complexes, which may contain additional components, and function at the cell surface or extracellularly, suggests as yet unexplored functions of these enzymes. Here we review how studies of basic biological processes in the mouse models of three lysosomal storage disorders, galactosialidosis, sialidosis, and GM1-gangliosidosis, revealed new and unexpected roles for the three respective affected enzymes, Ppca, Neu1, and β-Gal, that go beyond their canonical degradative activities. These findings have broadened our perspective on their functions and may pave the way for the development of new therapies for these lysosomal storage disorders.
Figures
References
-
- Galjart NJ, Gillemans N, Harris A, van der Horst GT, Verheijen FW, Galjaard H, d’Azzo A. Expression of cDNA encoding the human “protective protein” associated with lysosomal beta-galactosidase and neuraminidase: homology to yeast proteases. Cell. 1988;54(6):755–764. doi: 10.1016/S0092-8674(88)90999-3. - DOI - PubMed
-
- Pshezhetsky AV, Potier M. Association of N-acetylgalactosamine-6-sulfate sulfatase with the multienzyme lysosomal complex of beta-galactosidase, cathepsin A, and neuraminidase. Possible implication for intralysosomal catabolism of keratan sulfate. J Biol Chem. 1996;271(45):28359–28365. doi: 10.1074/jbc.271.45.28359. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
