

Evolutionary and structural analyses of mammalian haloacid dehalogenase-type phosphatases AUM and chronophin provide insight into the basis of their different substrate specificities
- PMID: 24338473
- PMCID: PMC3916544
- DOI: 10.1074/jbc.M113.503359
Evolutionary and structural analyses of mammalian haloacid dehalogenase-type phosphatases AUM and chronophin provide insight into the basis of their different substrate specificities
Abstract
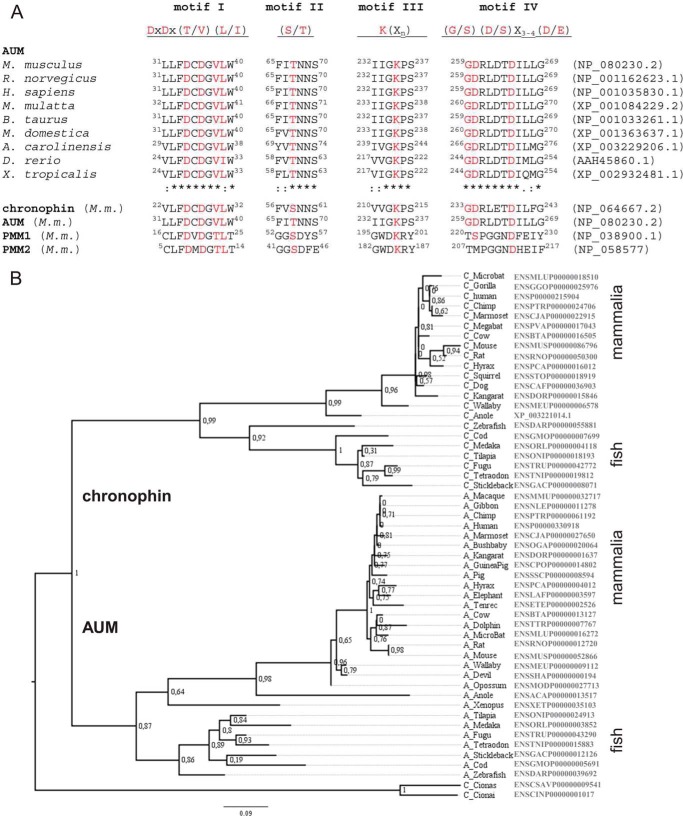
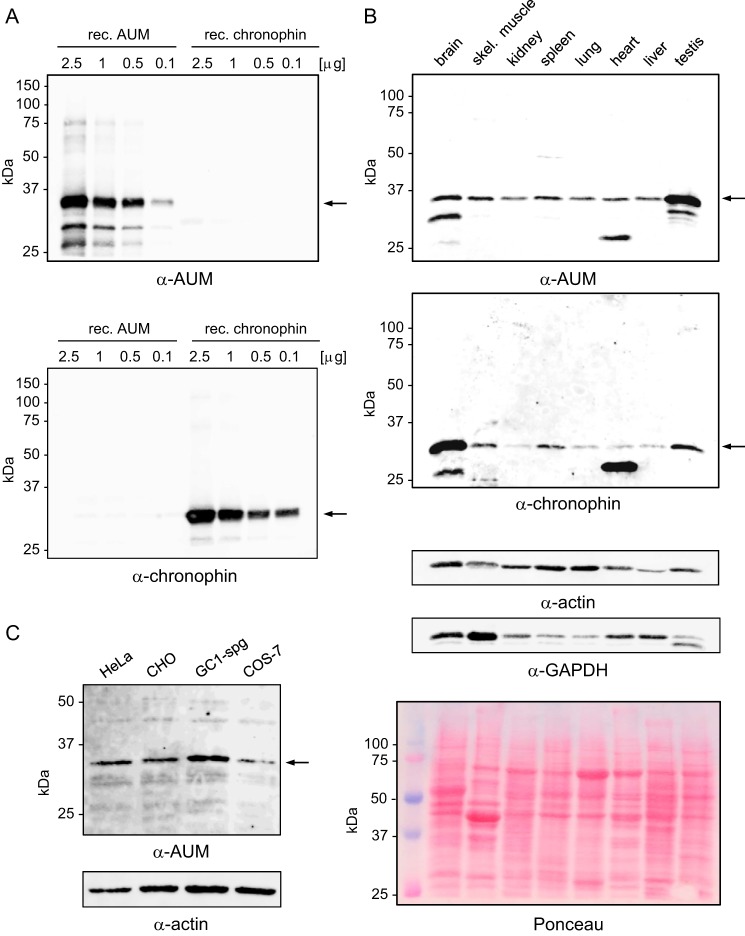
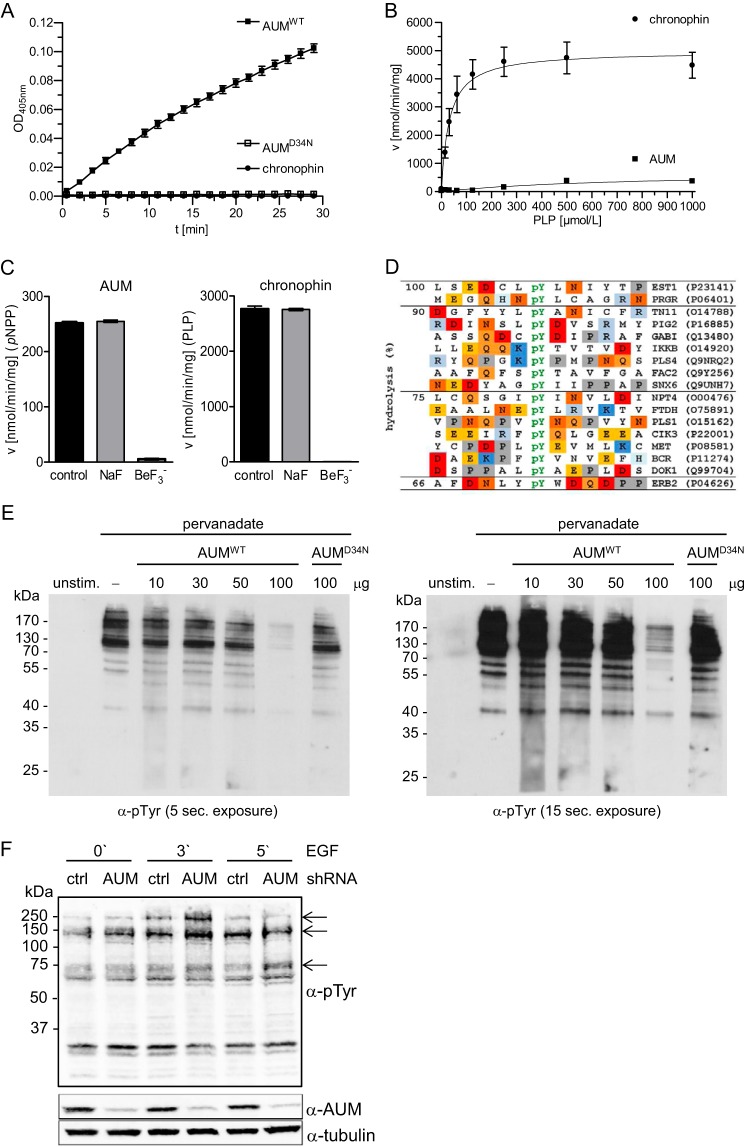
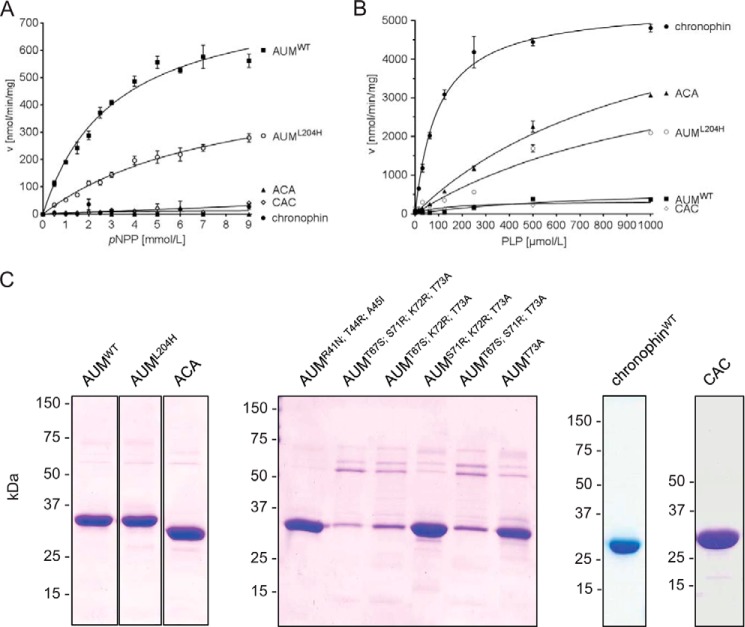
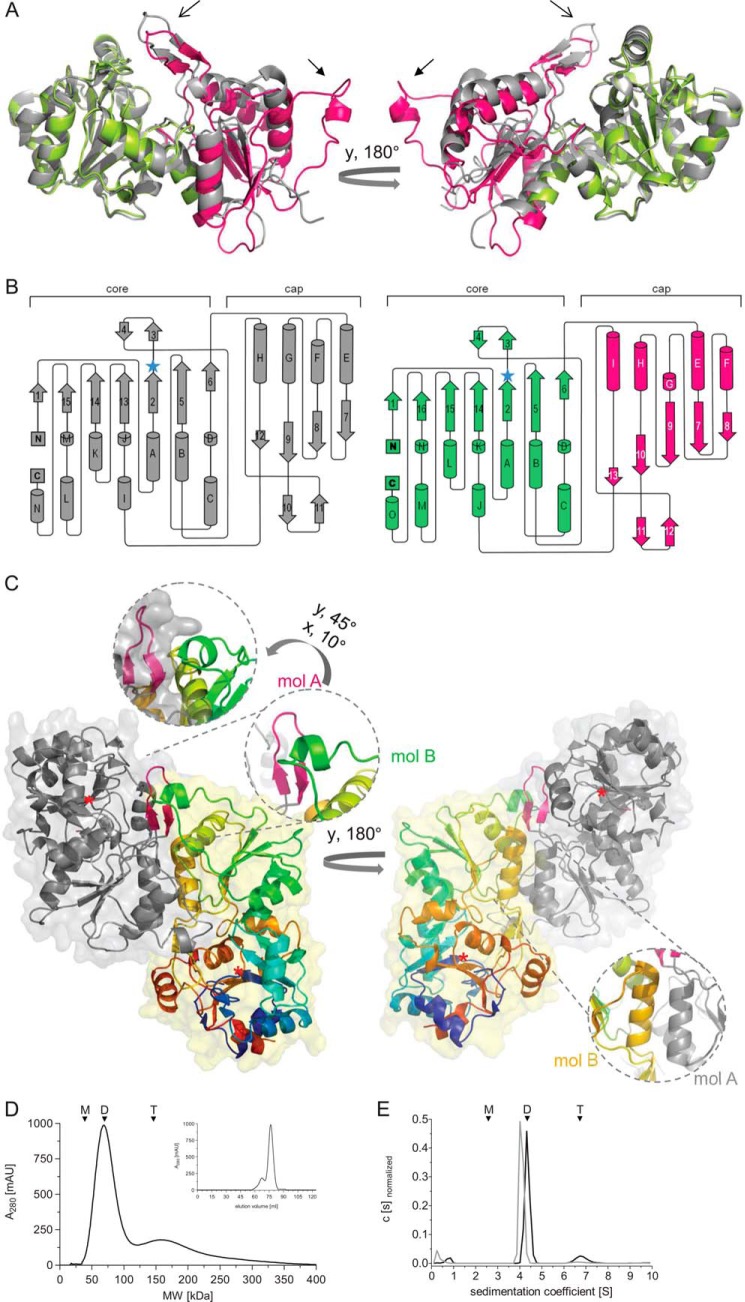
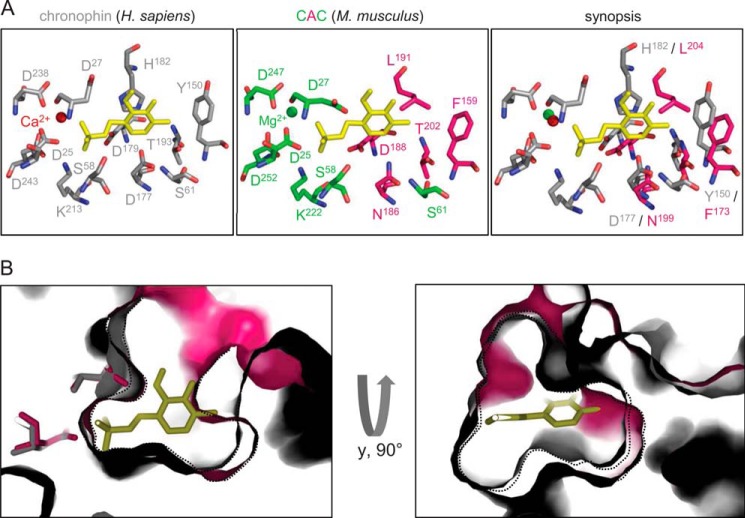
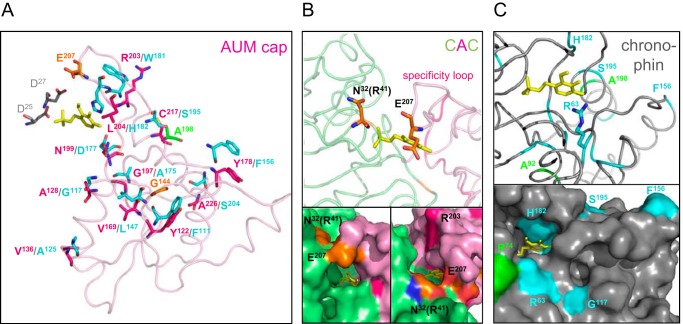
Mammalian haloacid dehalogenase (HAD)-type phosphatases are an emerging family of phosphatases with important functions in physiology and disease, yet little is known about the basis of their substrate specificity. Here, we characterize a previously unexplored HAD family member (gene annotation, phosphoglycolate phosphatase), which we termed AUM, for aspartate-based, ubiquitous, Mg(2+)-dependent phosphatase. AUM is a tyrosine-specific paralog of the serine/threonine-specific protein and pyridoxal 5'-phosphate-directed HAD phosphatase chronophin. Comparative evolutionary and biochemical analyses reveal that a single, differently conserved residue in the cap domain of either AUM or chronophin is crucial for phosphatase specificity. We have solved the x-ray crystal structure of the AUM cap fused to the catalytic core of chronophin to 2.65 Å resolution and present a detailed view of the catalytic clefts of AUM and chronophin that explains their substrate preferences. Our findings identify a small number of cap domain residues that encode the different substrate specificities of AUM and chronophin.
Keywords: Haloacid Dehalogenase Phosphatase; Molecular Evolution; Phosphatase Substrate Specificity; Phosphoglycolate Phosphatase; Pyridoxal Phosphate; Serine/Threonine Protein Phosphatase; Tyrosine-Protein Phosphatase (Tyrosine Phosphatase); X-ray Crystallography.
Figures
References
-
- Aravind L., Koonin E. V. (1998) The HD domain defines a new superfamily of metal-dependent phosphohydrolases. Trends Biochem. Sci. 23, 469–472 - PubMed
-
- Burroughs A. M., Allen K. N., Dunaway-Mariano D., Aravind L. (2006) Evolutionary genomics of the HAD superfamily: understanding the structural adaptations and catalytic diversity in a superfamily of phosphoesterases and allied enzymes. J. Mol. Biol. 361, 1003–1034 - PubMed
-
- Almo S. C., Bonanno J. B., Sauder J. M., Emtage S., Dilorenzo T. P., Malashkevich V., Wasserman S. R., Swaminathan S., Eswaramoorthy S., Agarwal R., Kumaran D., Madegowda M., Ragumani S., Patskovsky Y., Alvarado J., Ramagopal U. A., Faber-Barata J., Chance M. R., Sali A., Fiser A., Zhang Z. Y., Lawrence D. S., Burley S. K. (2007) Structural genomics of protein phosphatases. J. Struct. Funct. Genomics 8, 121–140 - PMC - PubMed
-
- Possemato R., Marks K. M., Shaul Y. D., Pacold M. E., Kim D., Birsoy K., Sethumadhavan S., Woo H. K., Jang H. G., Jha A. K., Chen W. W., Barrett F. G., Stransky N., Tsun Z. Y., Cowley G. S., Barretina J., Kalaany N. Y., Hsu P. P., Ottina K., Chan A. M., Yuan B., Garraway L. A., Root D. E., Mino-Kenudson M., Brachtel E. F., Driggers E. M., Sabatini D. M. (2011) Functional genomics reveal that the serine synthesis pathway is essential in breast cancer. Nature 476, 346–350 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
