

Strand-specific recognition of DNA damages by XPD provides insights into nucleotide excision repair substrate versatility
- PMID: 24338567
- PMCID: PMC3916561
- DOI: 10.1074/jbc.M113.523001
Strand-specific recognition of DNA damages by XPD provides insights into nucleotide excision repair substrate versatility
Abstract
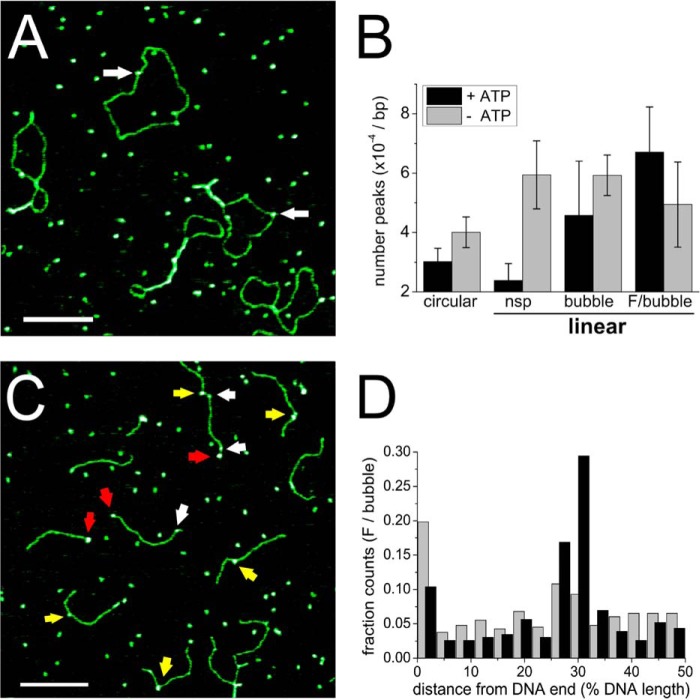
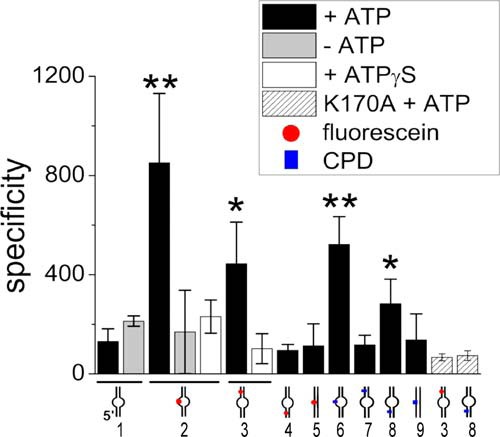
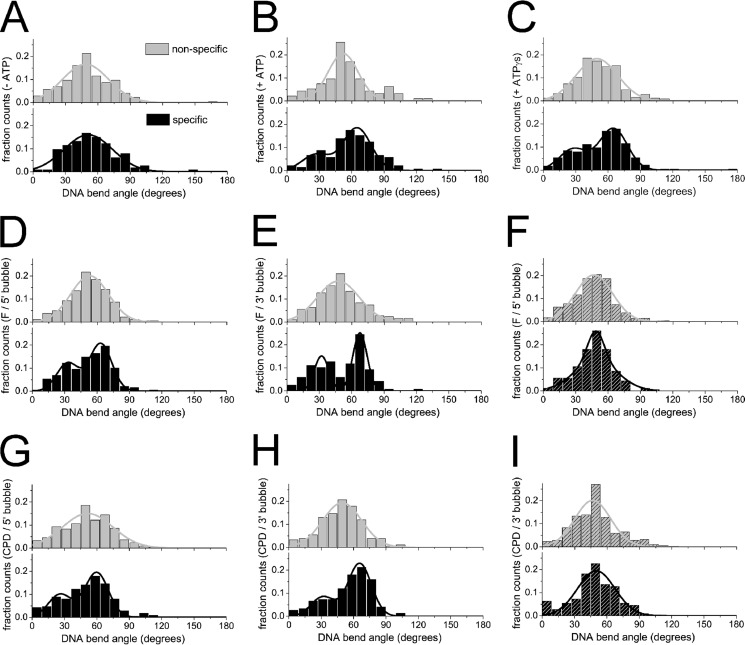
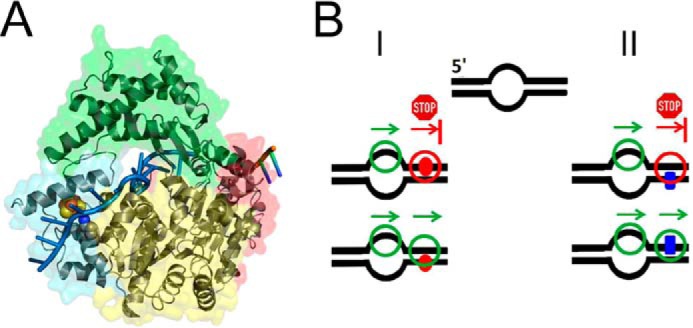
Recognition and removal of DNA damages is essential for cellular and organismal viability. Nucleotide excision repair (NER) is the sole mechanism in humans for the repair of carcinogenic UV irradiation-induced photoproducts in the DNA, such as cyclobutane pyrimidine dimers. The broad substrate versatility of NER further includes, among others, various bulky DNA adducts. It has been proposed that the 5'-3' helicase XPD (xeroderma pigmentosum group D) protein plays a decisive role in damage verification. However, despite recent advances such as the identification of a DNA-binding channel and central pore in the protein, through which the DNA is threaded, as well as a dedicated lesion recognition pocket near the pore, the exact process of target site recognition and verification in eukaryotic NER still remained elusive. Our single molecule analysis by atomic force microscopy reveals for the first time that XPD utilizes different recognition strategies to verify structurally diverse lesions. Bulky fluorescein damage is preferentially detected on the translocated strand, whereas the opposite strand preference is observed for a cyclobutane pyrimidine dimer lesion. Both states, however, lead to similar conformational changes in the resulting specific complexes, indicating a merge to a "final" verification state, which may then trigger the recruitment of further NER proteins.
Keywords: Atomic Force Microscopy; DNA Damage; DNA Helicase; DNA Lesion Recognition; DNA Repair; Protein-DNA Interactions; Single Molecule Biophysics.
Figures
Similar articles
-
Mechanism of DNA loading by the DNA repair helicase XPD.Nucleic Acids Res. 2016 Apr 7;44(6):2806-15. doi: 10.1093/nar/gkw102. Epub 2016 Feb 20. Nucleic Acids Res. 2016. PMID: 26896802 Free PMC article.
-
Strand- and site-specific DNA lesion demarcation by the xeroderma pigmentosum group D helicase.Proc Natl Acad Sci U S A. 2010 Oct 12;107(41):17545-50. doi: 10.1073/pnas.1004339107. Epub 2010 Sep 27. Proc Natl Acad Sci U S A. 2010. PMID: 20876134 Free PMC article.
-
The helicase XPD unwinds bubble structures and is not stalled by DNA lesions removed by the nucleotide excision repair pathway.Nucleic Acids Res. 2010 Jan;38(3):931-41. doi: 10.1093/nar/gkp1058. Epub 2009 Nov 20. Nucleic Acids Res. 2010. PMID: 19933257 Free PMC article.
-
The XPD helicase: XPanDing archaeal XPD structures to get a grip on human DNA repair.Biol Chem. 2010 Jul;391(7):761-5. doi: 10.1515/BC.2010.076. Biol Chem. 2010. PMID: 20482310 Review.
-
[Nucleotide excision repair in mammalia: mechanism of a primary damage recognition].Mol Biol (Mosk). 2008 Jan-Feb;42(1):24-31. Mol Biol (Mosk). 2008. PMID: 18389616 Review. Russian.
Cited by
-
Lesion search and recognition by thymine DNA glycosylase revealed by single molecule imaging.Nucleic Acids Res. 2015 Mar 11;43(5):2716-29. doi: 10.1093/nar/gkv139. Epub 2015 Feb 24. Nucleic Acids Res. 2015. PMID: 25712093 Free PMC article.
-
Close encounters for the first time: Helicase interactions with DNA damage.DNA Repair (Amst). 2015 Sep;33:43-59. doi: 10.1016/j.dnarep.2015.06.003. Epub 2015 Jun 16. DNA Repair (Amst). 2015. PMID: 26160335 Free PMC article. Review.
-
Transfer RNA methyltransferases from Thermoplasma acidophilum, a thermoacidophilic archaeon.Int J Mol Sci. 2014 Dec 23;16(1):91-113. doi: 10.3390/ijms16010091. Int J Mol Sci. 2014. PMID: 25546389 Free PMC article.
-
RecQ and Fe-S helicases have unique roles in DNA metabolism dictated by their unwinding directionality, substrate specificity, and protein interactions.Biochem Soc Trans. 2018 Feb 19;46(1):77-95. doi: 10.1042/BST20170044. Epub 2017 Dec 22. Biochem Soc Trans. 2018. PMID: 29273621 Free PMC article. Review.
-
Mechanistic and biological considerations of oxidatively damaged DNA for helicase-dependent pathways of nucleic acid metabolism.Free Radic Biol Med. 2017 Jun;107:245-257. doi: 10.1016/j.freeradbiomed.2016.11.022. Epub 2016 Nov 22. Free Radic Biol Med. 2017. PMID: 27884703 Free PMC article. Review.
References
-
- Friedberg E. C. (2003) DNA damage and repair. Nature 421, 436–440 - PubMed
-
- Lindahl T., Wood R. D. (1999) Quality control by DNA repair. Science 286, 1897–1905 - PubMed
-
- Hoeijmakers J. H. (2001) Genome maintenance mechanisms for preventing cancer. Nature 411, 366–374 - PubMed
-
- Friedberg E. C. (2001) How nucleotide excision repair protects against cancer. Nat. Rev. Cancer 1, 22–33 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
