

Photoaffinity labeling the propofol binding site in GLIC
- PMID: 24341978
- PMCID: PMC3935609
- DOI: 10.1021/bi401492k
Photoaffinity labeling the propofol binding site in GLIC
Abstract
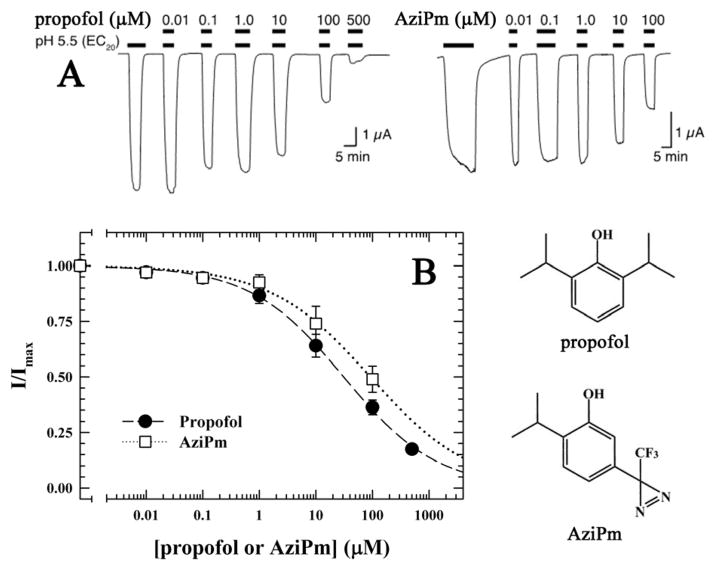
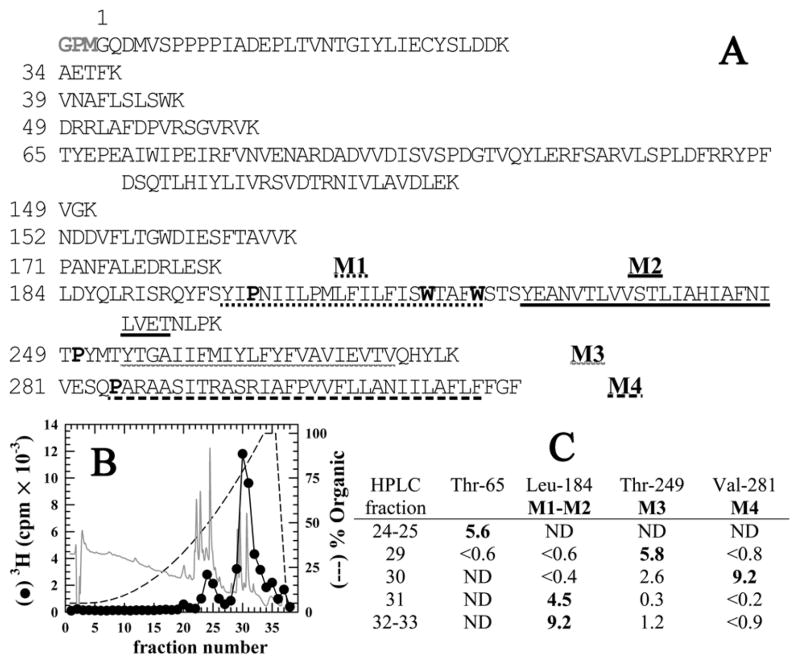
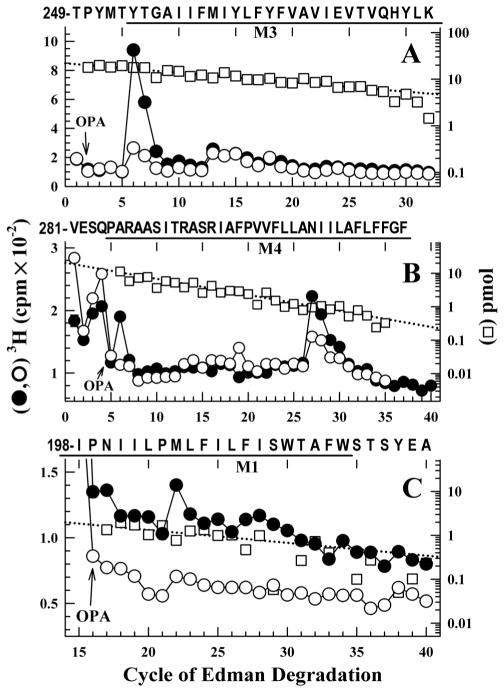
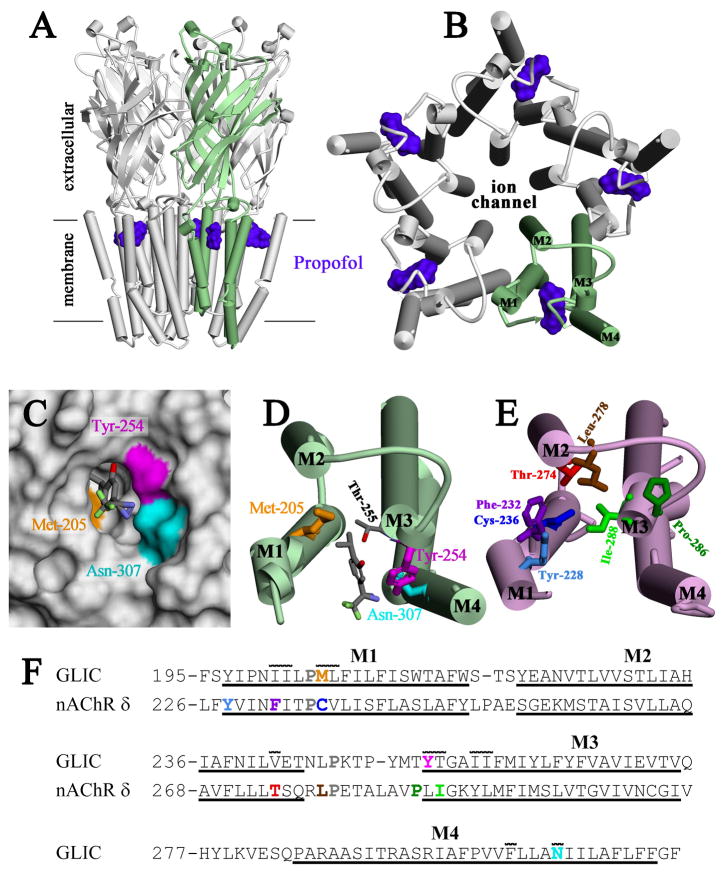
Propofol, an intravenous general anesthetic, produces many of its anesthetic effects in vivo by potentiating the responses of GABA type A receptors (GABAAR), members of the superfamily of pentameric ligand-gated ion channels (pLGICs) that contain anion-selective channels. Propofol also inhibits pLGICs containing cation-selective channels, including nicotinic acetylcholine receptors and GLIC, a prokaryotic proton-gated homologue from Gloeobacter violaceus . In the structure of GLIC cocrystallized with propofol at pH 4 (presumed open/desensitized states), propofol was localized to an intrasubunit pocket at the extracellular end of the transmembrane domain within the bundle of transmembrane α-helices (Nury, H, et al. (2011) Nature 469, 428-431). To identify propofol binding sites in GLIC in solution, we used a recently developed photoreactive propofol analogue (2-isopropyl-5-[3-(trifluoromethyl)-3H-diazirin-3-yl]phenol or AziPm) that acts as an anesthetic in vivo and potentiates GABAAR in vitro. For GLIC expressed in Xenopus oocytes, propofol and AziPm inhibited current responses at pH 5.5 (EC20) with IC50 values of 20 and 50 μM, respectively. When [(3)H]AziPm (7 μM) was used to photolabel detergent-solubilized, affinity-purified GLIC at pH 4.4, protein microsequencing identified propofol-inhibitable photolabeling of three residues in the GLIC transmembrane domain: Met-205, Tyr-254, and Asn-307 in the M1, M3, and M4 transmembrane helices, respectively. Thus, for GLIC in solution, propofol and AziPm bind competitively to a site in proximity to these residues, which, in the GLIC crystal structure, are in contact with the propofol bound in the intrasubunit pocket.
Conflict of interest statement
Figures
References
-
- Krasowski MD, Jenkins A, Flood P, Kung AY, Hopfinger AJ, Harrison NL. General anesthetic potencies of a series of propofol analogs correlate with potency for potentiation of gamma-aminobutyric acid (GABA) current at the GABA(A) receptor but not with lipid solubility. J Pharmacol Exp Ther. 2001;297:338–351. - PubMed
-
- Rudolph U, Antkowiak B. Molecular and neuronal substrates for general anaesthetics. Nat Rev Neurosci. 2004;5:709–720. - PubMed
-
- Franks NP. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nat Rev Neurosci. 2008;9:370–386. - PubMed
-
- Dilger JP, Vidal AM, Mody HI, Liu Y. Evidence for direct actions of general anesthetics on an ion channel protein - A new look at a unified mechanism of action. Anesthesiology. 1994;81:431–442. - PubMed
-
- Violet JM, Downie DL, Nakisa RC, Lieb WR, Franks NP. Differential sensitivities of mammalian neuronal and muscle nicotinic acetylcholine receptors to general anesthetics. Anesthesiology. 1997;86:866–874. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
