

Review
doi: 10.1038/nrg3623.
Epub 2013 Dec 17.
Dynamic regulation of transcriptional states by chromatin and transcription factors
Affiliations
- PMID: 24342920
- PMCID: PMC6322398
- DOI: 10.1038/nrg3623
Item in Clipboard
Review
Dynamic regulation of transcriptional states by chromatin and transcription factors
Nat Rev Genet.
2014 Feb.
Abstract
The interaction of regulatory proteins with the complex nucleoprotein structures that are found in mammalian cells involves chromatin reorganization at multiple levels. Mechanisms that support these transitions are complex on many timescales, which range from milliseconds to minutes or hours. In this Review, we discuss emerging concepts regarding the function of regulatory elements in living cells. We also explore the involvement of these dynamic and stochastic processes in the evolution of fluctuating transcriptional activity states that are now commonly reported in eukaryotic systems.
Conflict of interest statement
Competing interests statement
The authors declare no competing interests.
Figures
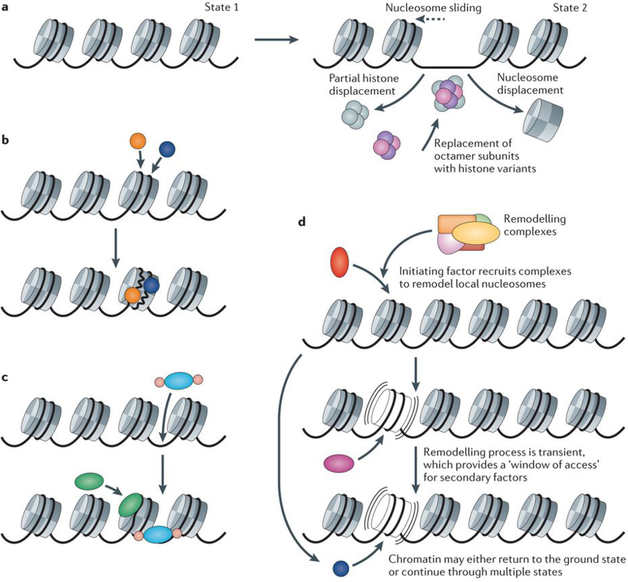
a | Most current models that invoke either ATP-dependent remodelling or histone chaperone action envisage a factor-dependent conversion of one static chromatin state to an alternative state. Possible mechanisms for increased local access include nucleosome sliding (shown by the dashed arrow), nucleosome displacement, partial histone displacement and replacement of octamer subunits with histone variants (shown in pink and purple)–. b | One of the classic mechanisms for factor penetration of closed chromatin structures is cooperative nucleosome attack. Although one protein may not be able to dislodge or reorganize local nucleosome structures, it has been suggested that two factors (shown in orange and blue) that act in tandem cooperatively generate sufficient free energy of binding to overcome the many histone–DNA contacts in the nucleosome, which leads to a new state with static factor binding. c | In another mechanism, pioneer proteins are argued to have special properties that allow their interaction with closed chromatin. For the forkhead box protein A (FOXA) and FOXO families of proteins (shown in blue and pink, respectively), their structural similarity with histone H1 may lead to ‘wedging’ of DNA–histone contacts, which allows secondary access of other transcription factors (shown in green). Neither of these concepts invoke a necessary role for ATP-dependent remodelling complexes in the generation of altered nucleosome states. d | An alternative view suggests that the averaged states that are observed in cell populations result from a highly dynamic interconversion between multiple local chromatin states. A recent example of this concept is the assisted loading model for nuclear receptor action, in which receptors recruit multiple remodelling complexes to induce local chromatin conformations that have short lifetimes. These modulated states provide transient ‘windows of access’, during which secondary factors (shown in red, pink and blue) gain access to their binding sites. This process may either continue in a linear mode, which generates a series of altered chromatin states, or involve continuous cycling.
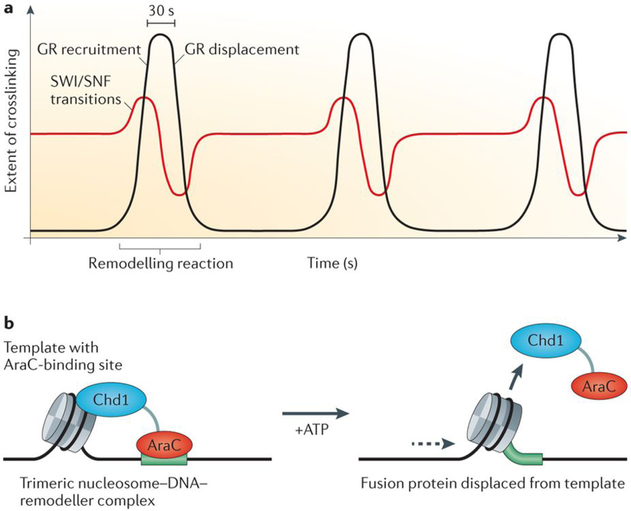
a | The extensive reorganization of nucleosome structures during ATP-dependent remodelling can induce unexpected consequences for transcription factor binding. Glucocorticoid receptor (GR) binding during receptor-induced nucleosome B remodelling at the mouse mammary tumour virus promoter by the SWI/SNF complex was studied in vitro, following real-time factor–template interactions with high-speed UV laser crosslinking. Pulses of GR recruitment and displacement (shown in black) were found to require ATP-dependent remodelling and to correlate with transient changes in BRG1–template crosslinking (shown in red). It was concluded that the receptor was actively ejected from the template during remodelling. b | One study characterized transcription factor binding during nucleosome sliding that is induced by a fusion protein between the yeast chromodomain-containing protein 1 (Chd1) and the DNA-binding domain of the Escherichia coli arabinose operon regulatory protein (AraC). This synthetic system allows tethering of the Chd1 remodelling protein domain to a specific binding site (shown in green), which is mediated by the AraC DNA-binding domain. Sliding of a positioned nucleosome to the centre of the DNA template (shown by the dashed arrow) results in an eviction of the fusion protein from the template by nucleosome invasion of the binding site. This phenomenon shows how factor mobility can result from the large-scale macromolecular reorganization that occurs during chromatin remodelling.
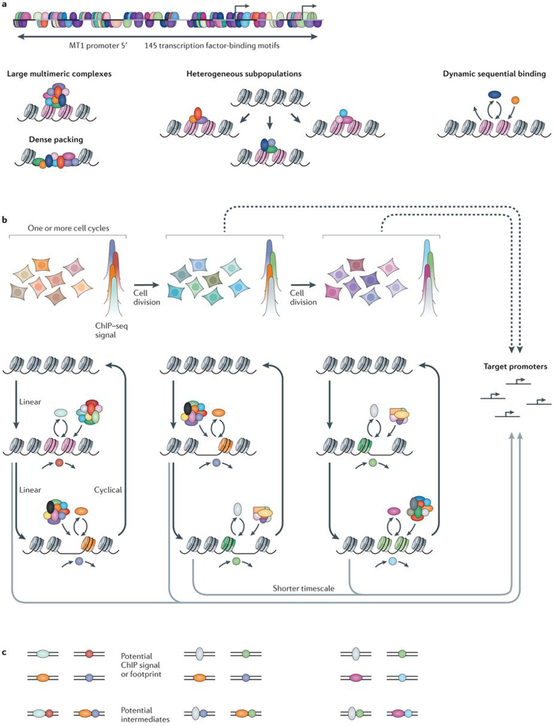
a | Mammalian promoter and enhancer elements are densely packed with transcription factor-binding motifs. An analysis using the Genomatix software suite showed that the 756‑bp region upstream of the metallothionein 1 (MT1) promoter contains 145 motifs, many of which are overlapping. In addition, a chromatin immunoprecipitation followed by sequencing (ChIP–seq) analysis reveals numerous localized elements that bind to as many as 40–50 factors (both are shown by semicircles). This conundrumcan be resolved by three alternative paradigms: large, static, multimeric complexes with dense packing; heterogeneous subpopulations that are bound by different subsets of factors; or dynamic sequential binding models that envision multiple transient states. Pink nucleosomes represent regions of chromatin that are undergoing dynamic reorganization, which results in accessible or ‘open’ chromatin. b | A dynamic view of enhancer functionis shown. The association between transcription factorsand regulatory elements is usually detected by ChIP–seq analyses across large cell populations. These cell groupsare highly heterogeneous with respect to local chromatin states and factor occupancy; variation is indicated schematically in the cell population by different colours. For brevity, four putative binding factors are depicted at a generic enhancer and their ChIP–seq signals are shown, although many more are often detected as co-binding. In static models, it is assumed that these factors form long-lived, multiprotein complexes. However, many dynamic processes within individual cells contribute to the population-averaged signal. Multiple chromatin remodelling complexes are probably recruited by transcription factors (which are colour coded to the ChIP– seq signal) to these elements; it is not known whether these processes are linear and sequential or whether they are cyclical. As cell division occurs, the population average occupancy, as well as participation in the underlying dynamic events (that is, assisted loading (FIG. 1)), evolves. A common assumption is that static, long-lived enhancer states that are observed across the cell population interact over long ranges to target promoters as shown by the dashed arrows. In fact, the chromatin states with brief lifetimes that contribute to the population average may separately communicate with promoters on a much shorter timescale. c | The dynamic model predicts many intermediate states. Some states may have lifetimes that are sufficient to produce factor footprints in cell population experiments, whereas others are probably too short-livedto be detected, even by formaldehyde-based ChIP.
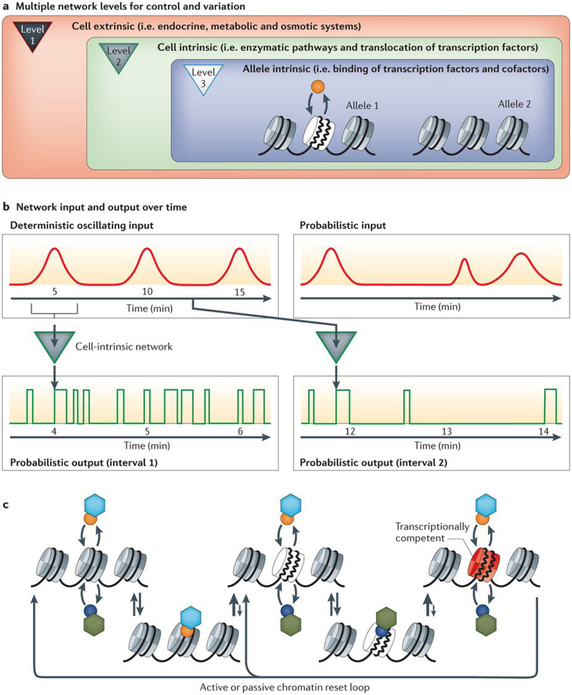
a | Dynamic events produce different temporal response patterns. Some of these responses — which range from intracellular signalling to transcriptional control — are highly reproducible and seem to be deterministic, whereas other responses are much more random and/or probabilistic in nature. Signalling network loops operate at the level of the organism (level 1), the individual cell (level 2) and the individual allele (level 3). Depending on specific configurations, these multiple levels can interact with one another to either increase or decrease the ordered activation of transcription. b | The top left panel shows a deterministic oscillating temporal signal that regulates a cell-intrinsic transcriptional control system. This stochastic signalling network may respond to these inputs either by increasing frequency of output signals during stimulation (bottom left panel) or by decreasing frequency in the absence of stimulation (bottom right panel). The top right panel indicates that the oscillating signal may be probabilistic rather than deterministic, which introduces further stochasticity in the system. c |The ‘hit-and-run’ transcriptional control model involves dynamic protein–protein interactions between transcription factors (hexagons) and co-regulators (circles), as well as protein–DNA interactions between these proteins and their target DNA. This stochastic process produces a chromatin template that cycles through multiple states, including a transcriptionally competent state (shown in red). This process may produce a deterministic linear series, or it may be partially reversible. The output of these dynamic states integrated over time produces transcriptional activation events that seem to be deterministic at the averaged whole-cell population level.
References
-
- Spitz F & Furlong EE Transcription factors: from enhancer binding to developmental control. Nature Rev. Genet 13, 613–626 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
