

Higher-order unfolding of satellite heterochromatin is a consistent and early event in cell senescence
- PMID: 24344186
- PMCID: PMC3871423
- DOI: 10.1083/jcb.201306073
Higher-order unfolding of satellite heterochromatin is a consistent and early event in cell senescence
Abstract
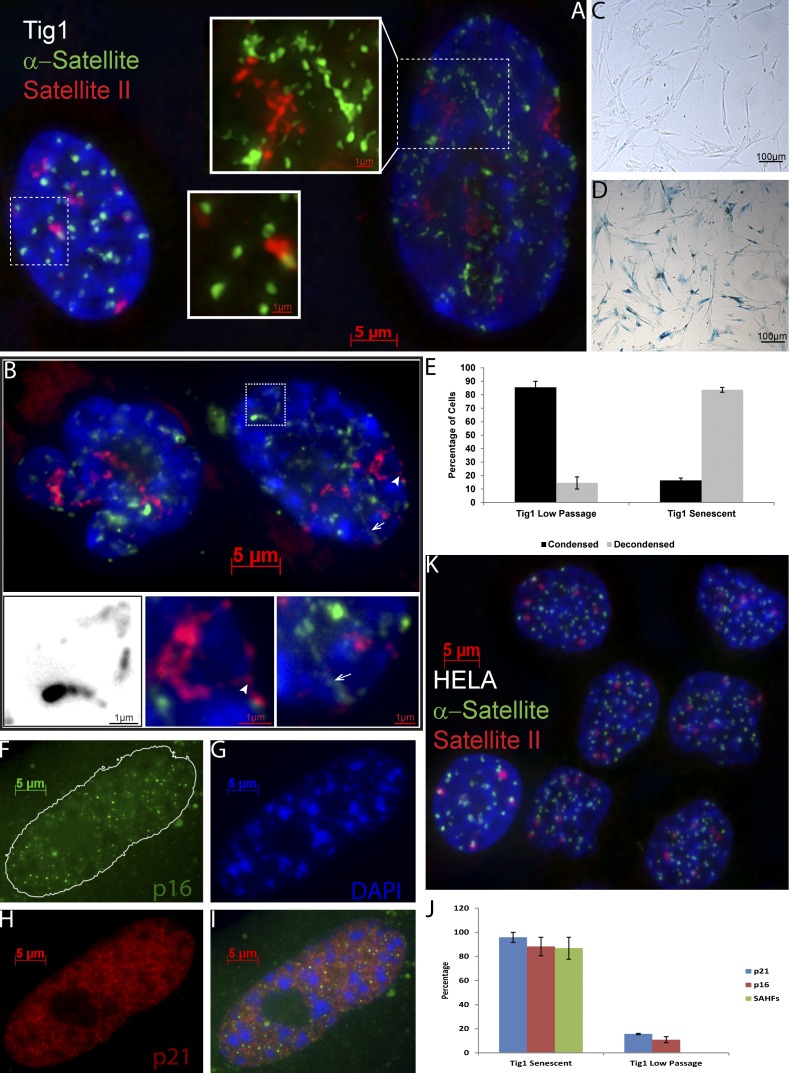
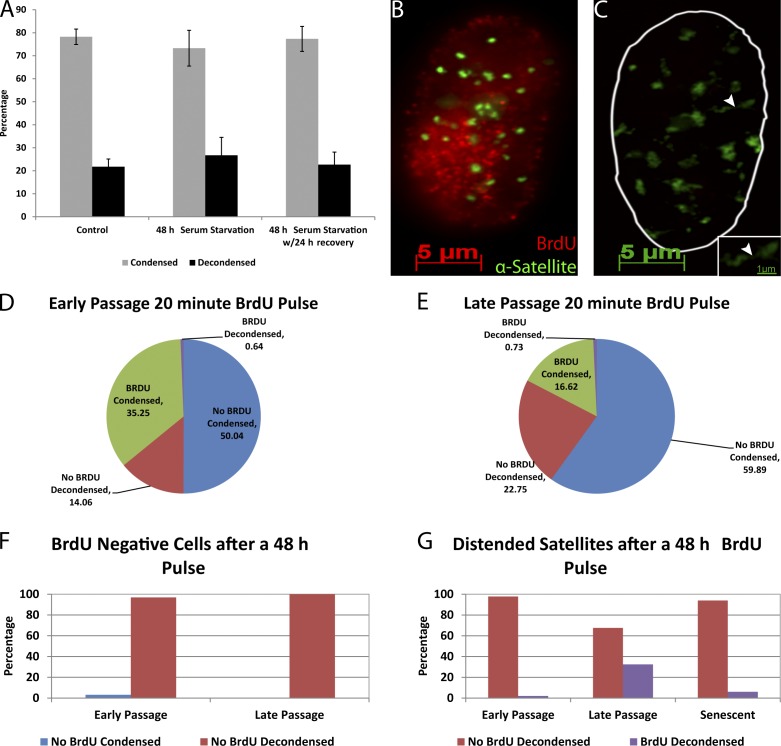
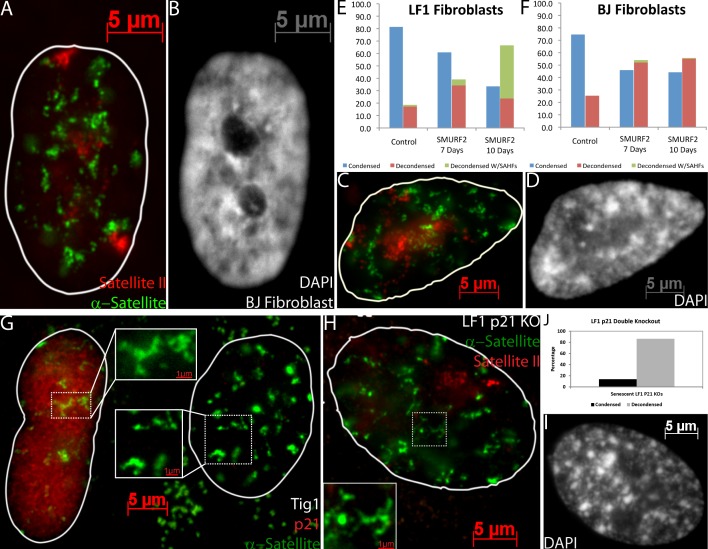
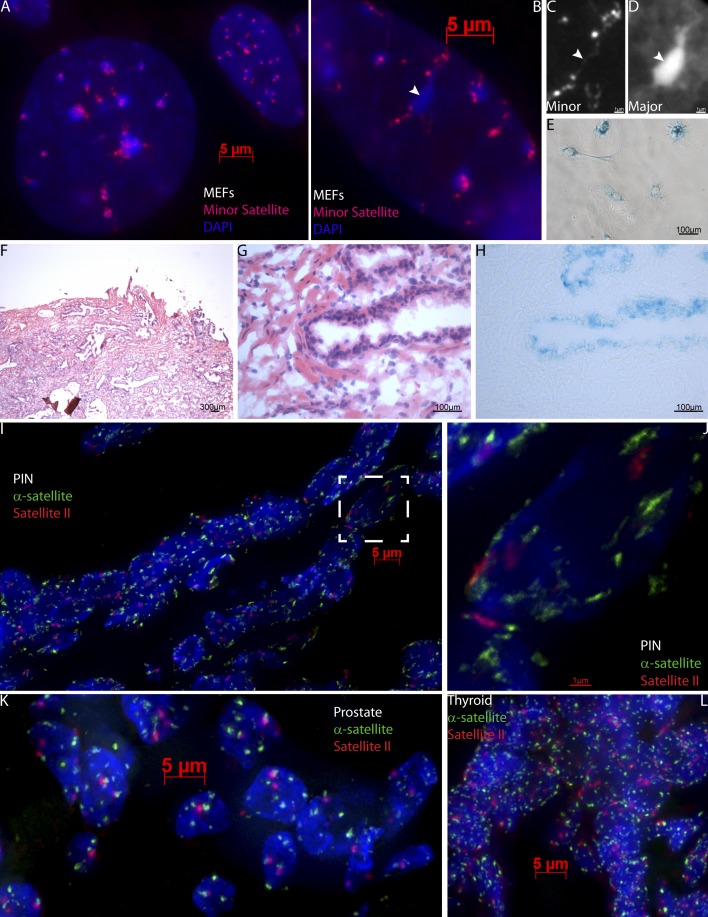
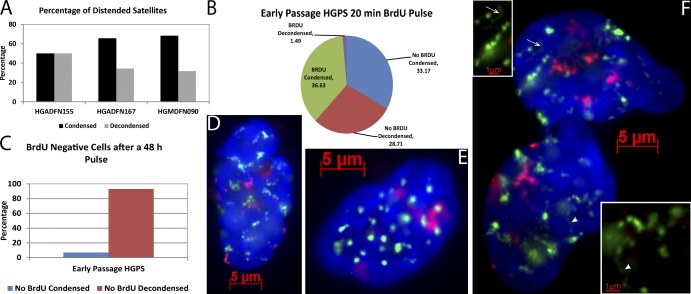
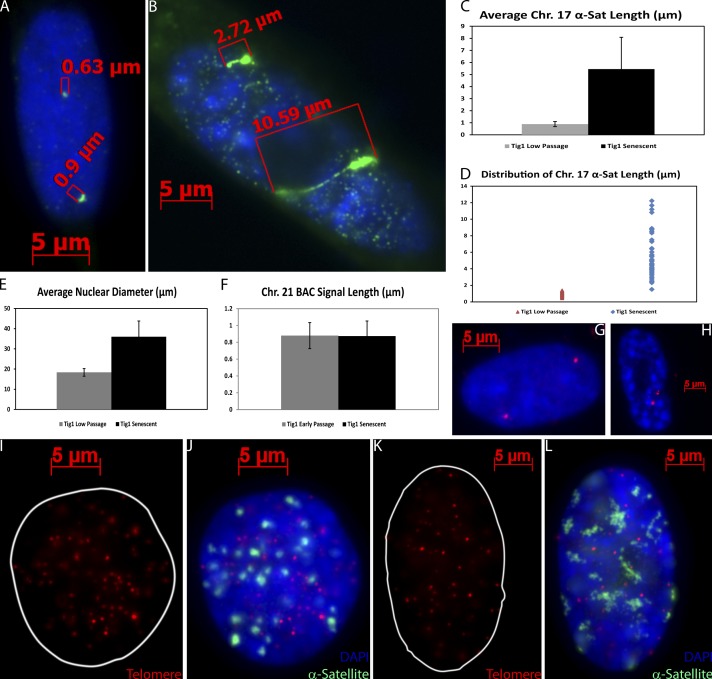
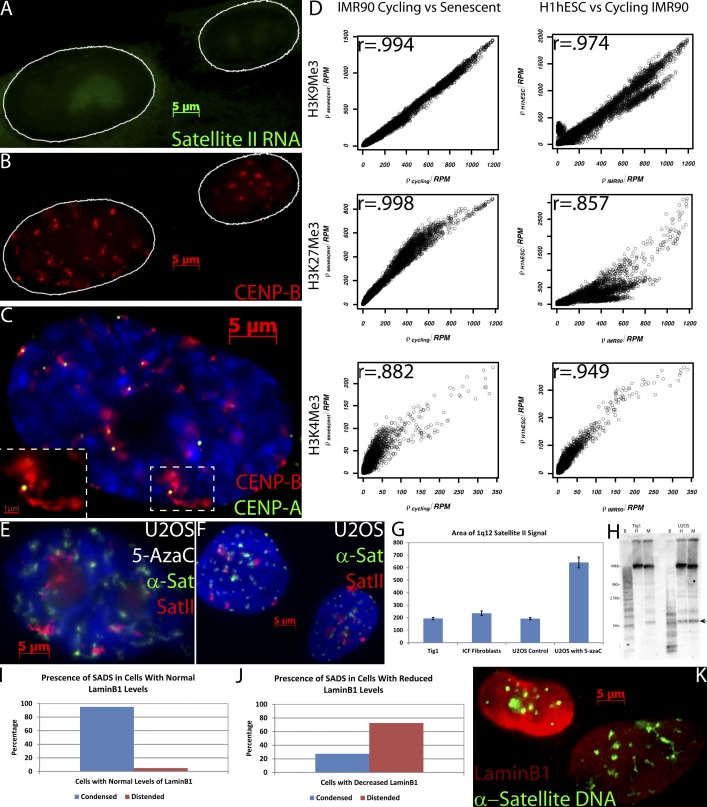
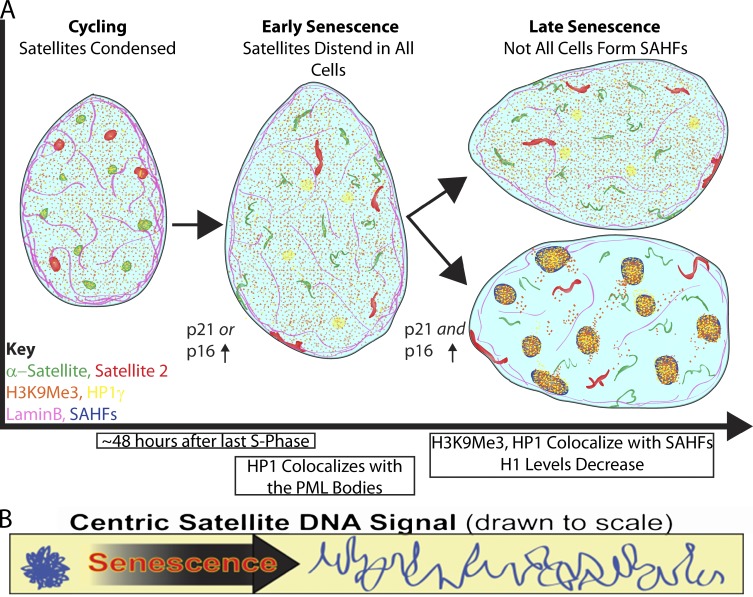
Epigenetic changes to chromatin are thought to be essential to cell senescence, which is key to tumorigenesis and aging. Although many studies focus on heterochromatin gain, this work demonstrates large-scale unraveling of peri/centromeric satellites, which occurs in all models of human and mouse senescence examined. This was not seen in cancer cells, except in a benign senescent tumor in vivo. Senescence-associated distension of satellites (SADS) occurs earlier and more consistently than heterochromatin foci formation, and SADS is not exclusive to either the p16 or p21 pathways. Because Hutchinson Guilford progeria syndrome patient cells do not form excess heterochromatin, the question remained whether or not proliferative arrest in this aging syndrome involved distinct epigenetic mechanisms. Here, we show that SADS provides a unifying event in both progeria and normal senescence. Additionally, SADS represents a novel, cytological-scale unfolding of chromatin, which is not concomitant with change to several canonical histone marks nor a result of DNA hypomethylation. Rather, SADS is likely mediated by changes to higher-order nuclear structural proteins, such as LaminB1.
Figures
References
-
- Chandra T., Kirschner K., Thuret J.-Y., Pope B.D., Ryba T., Newman S., Ahmed K., Samarajiwa S.A., Salama R., Carroll T., et al. 2012. Independence of repressive histone marks and chromatin compaction during senescent heterochromatic layer formation. Mol. Cell. 47:203–214 10.1016/j.molcel.2012.06.010 - DOI - PMC - PubMed
-
- De Cecco M., Criscione S.W., Peckham E.J., Hillenmeyer S., Hamm E.A., Manivannan J., Peterson A.L., Kreiling J.A., Neretti N., Sedivy J.M. 2013. Genomes of replicatively senescent cells undergo global epigenetic changes leading to gene silencing and activation of transposable elements. Aging Cell. 12:247–256 10.1111/acel.12047 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
