

Oligomerization transforms human APOBEC3G from an efficient enzyme to a slowly dissociating nucleic acid-binding protein
- PMID: 24345943
- PMCID: PMC3950479
- DOI: 10.1038/nchem.1795
Oligomerization transforms human APOBEC3G from an efficient enzyme to a slowly dissociating nucleic acid-binding protein
Abstract
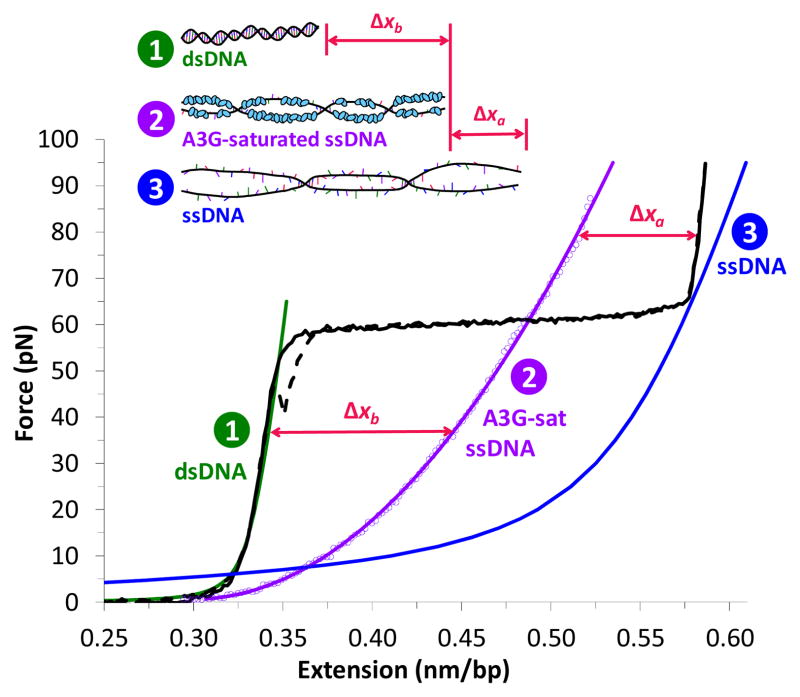
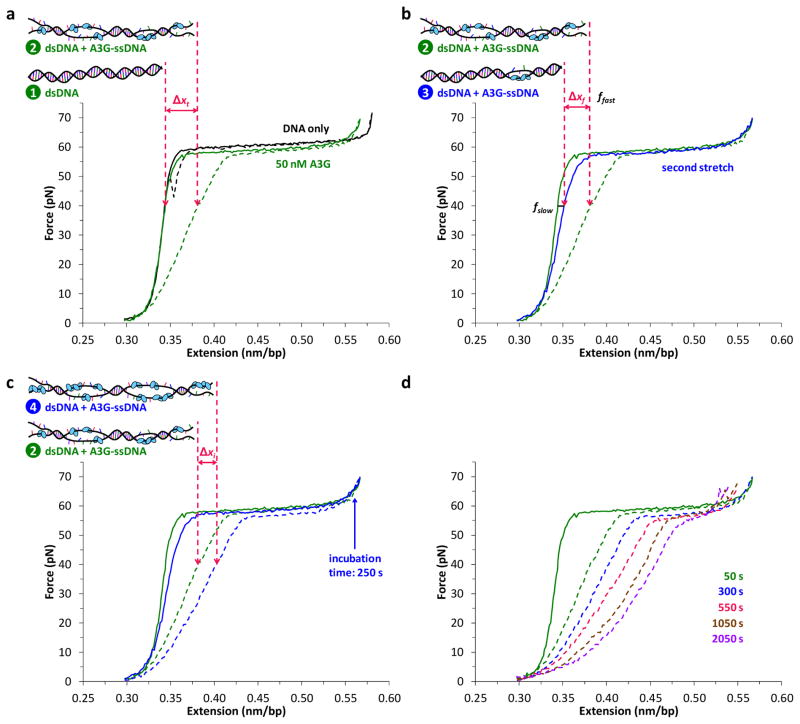
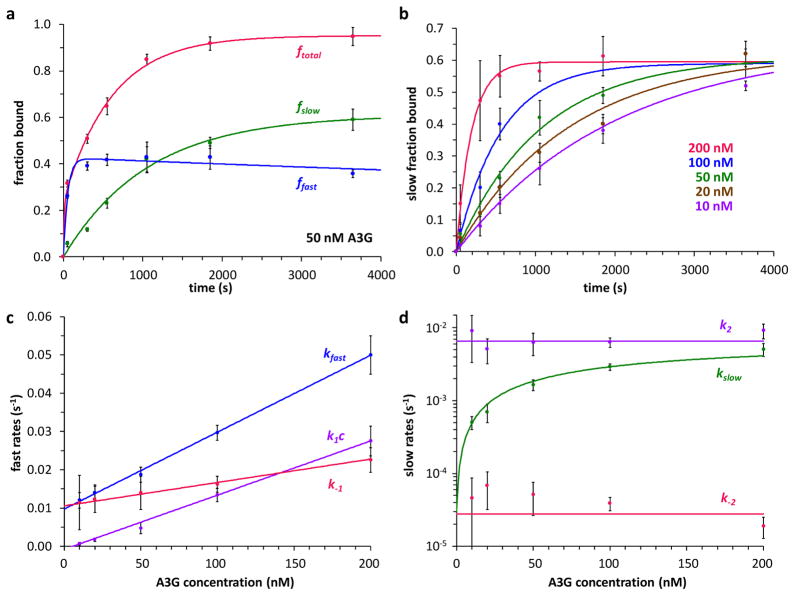
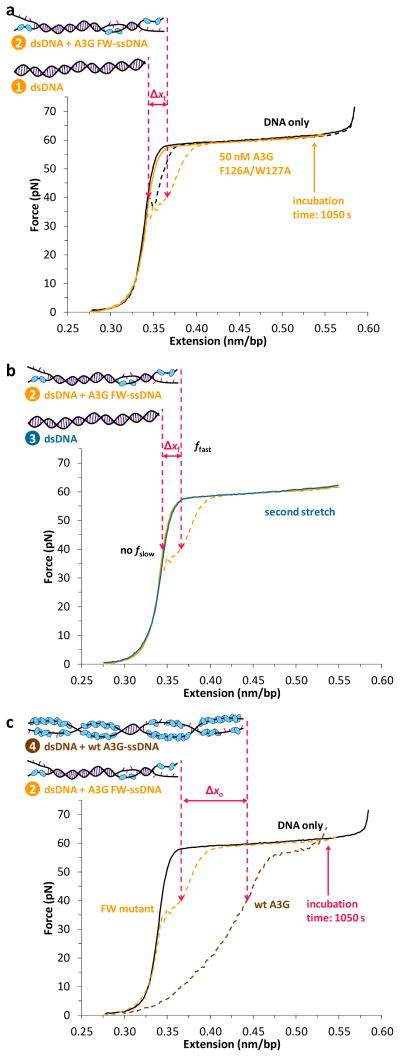
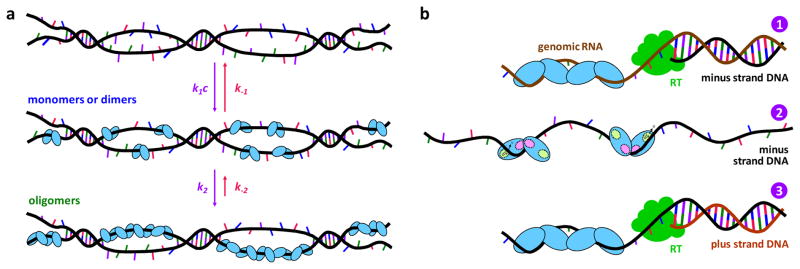
The human APOBEC3 proteins are a family of DNA-editing enzymes that play an important role in the innate immune response against retroviruses and retrotransposons. APOBEC3G is a member of this family that inhibits HIV-1 replication in the absence of the viral infectivity factor Vif. Inhibition of HIV replication occurs by both deamination of viral single-stranded DNA and a deamination-independent mechanism. Efficient deamination requires rapid binding to and dissociation from ssDNA. However, a relatively slow dissociation rate is required for the proposed deaminase-independent roadblock mechanism in which APOBEC3G binds the viral template strand and blocks reverse transcriptase-catalysed DNA elongation. Here, we show that APOBEC3G initially binds ssDNA with rapid on-off rates and subsequently converts to a slowly dissociating mode. In contrast, an oligomerization-deficient APOBEC3G mutant did not exhibit a slow off rate. We propose that catalytically active monomers or dimers slowly oligomerize on the viral genome and inhibit reverse transcription.
Figures
Comment in
-
Biophysical chemistry: Strength in numbers.Nat Chem. 2014 Jan;6(1):13-4. doi: 10.1038/nchem.1831. Nat Chem. 2014. PMID: 24345937 No abstract available.
References
-
- Harris RS, Liddament MT. Retroviral restriction by APOBEC proteins. Nat Rev Immunol. 2004;4(11):868–877. - PubMed
-
- Chiu YL, Greene WC. The APOBEC3 cytidine deaminases: An innate defensive network opposing exogenous retroviruses and endogenous retroelements. Annu Rev Immunol. 2008;26:317–353. - PubMed
-
- Sheehy AM, Gaddis NC, Choi JD, Malim MH. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature. 2002;418(6898):646–650. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
