

dcc orchestrates the development of the prefrontal cortex during adolescence and is altered in psychiatric patients
- PMID: 24346136
- PMCID: PMC4030324
- DOI: 10.1038/tp.2013.105
dcc orchestrates the development of the prefrontal cortex during adolescence and is altered in psychiatric patients
Abstract
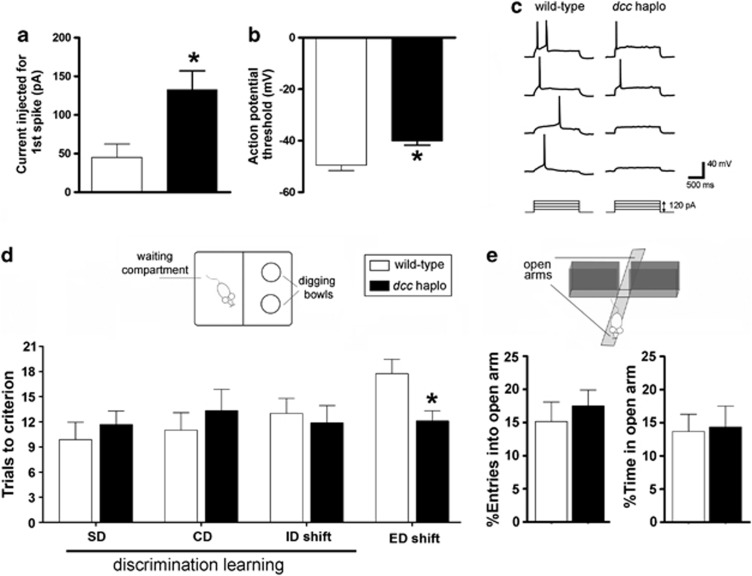
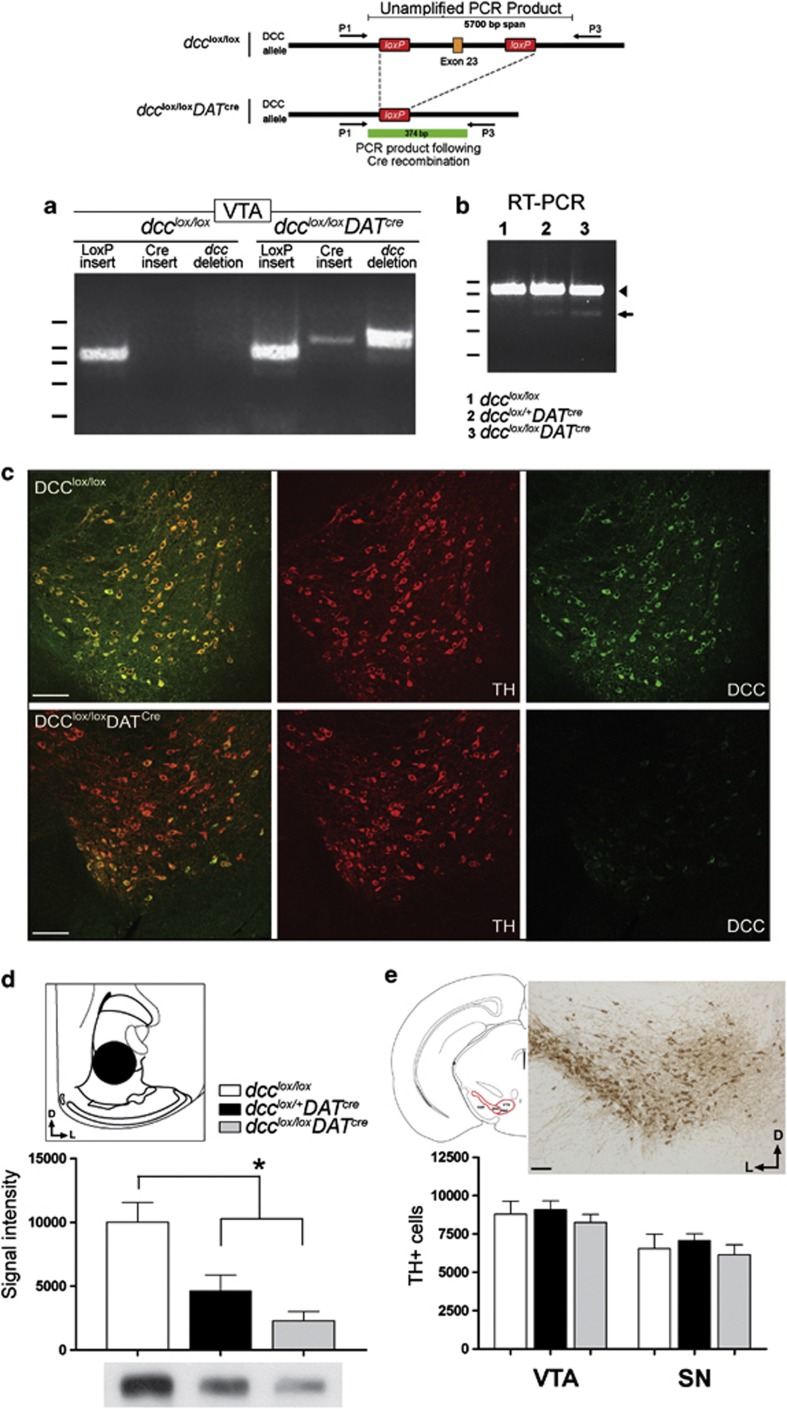
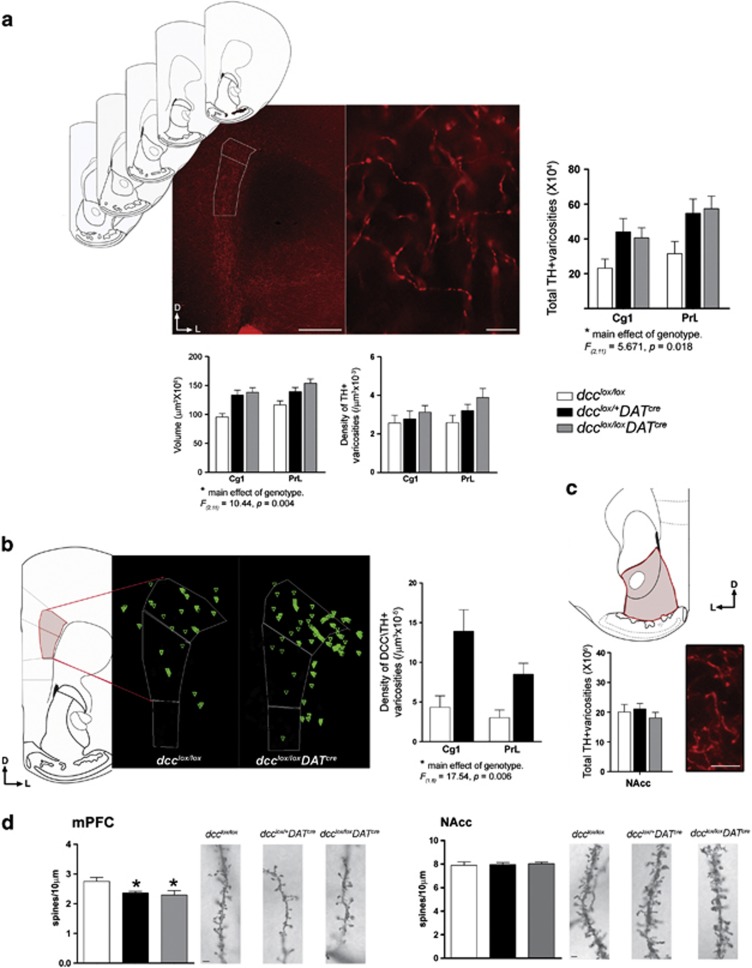
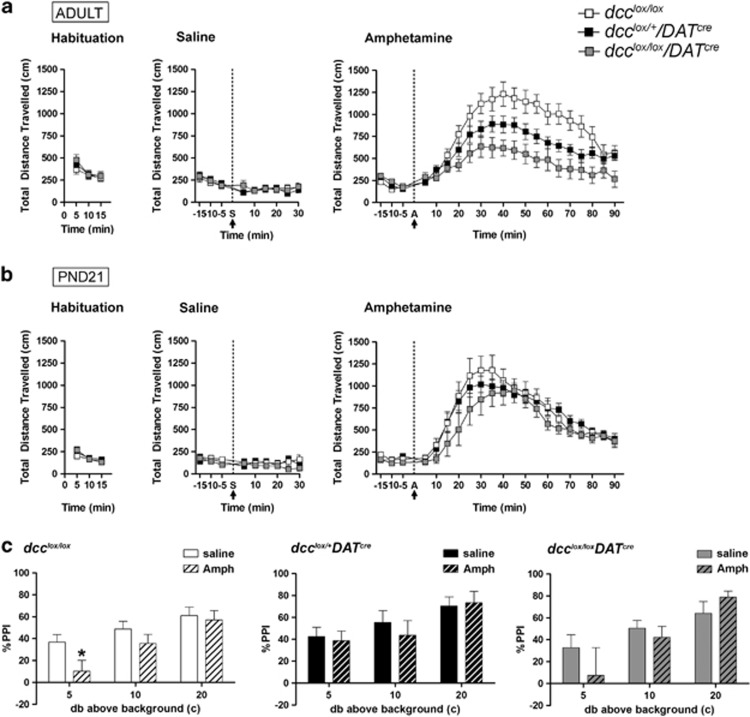
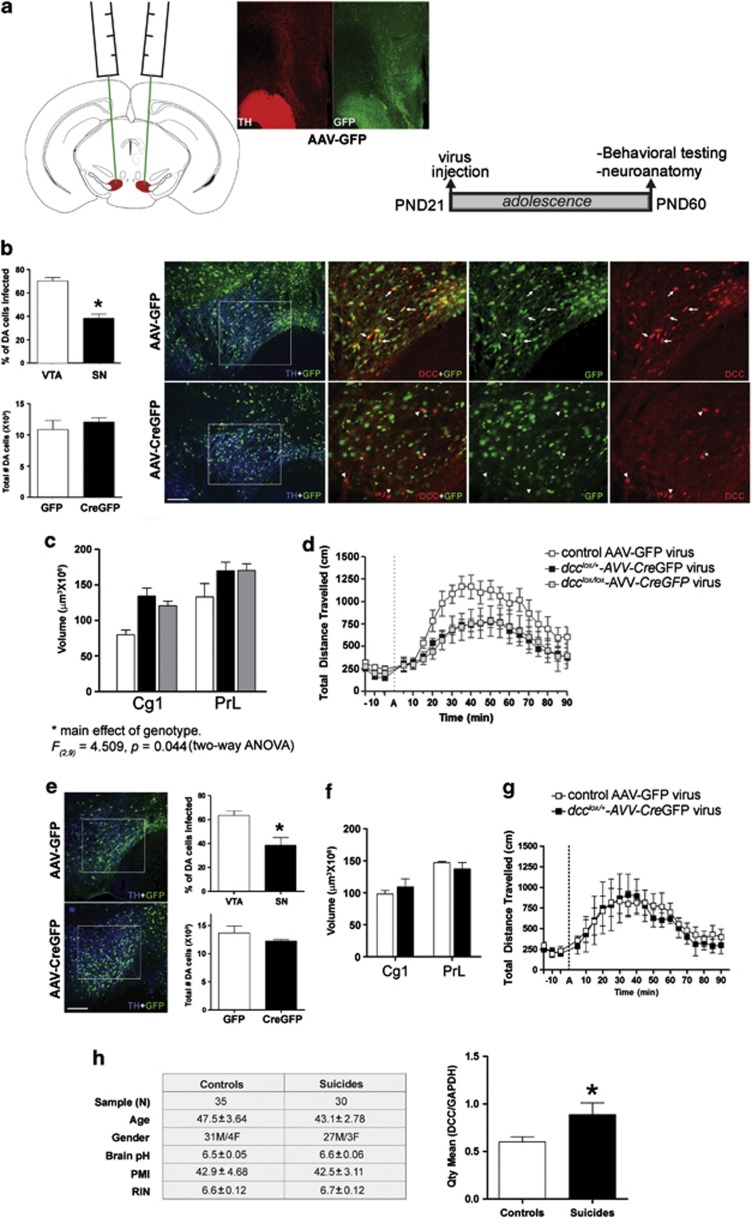
Adolescence is a period of heightened susceptibility to psychiatric disorders of medial prefrontal cortex (mPFC) dysfunction and cognitive impairment. mPFC dopamine (DA) projections reach maturity only in early adulthood, when their control over cognition becomes fully functional. The mechanisms governing this protracted and unique development are unknown. Here we identify dcc as the first DA neuron gene to regulate mPFC connectivity during adolescence and dissect the mechanisms involved. Reduction or loss of dcc from DA neurons by Cre-lox recombination increased mPFC DA innervation. Underlying this was the presence of ectopic DA fibers that normally innervate non-cortical targets. Altered DA input changed the anatomy and electrophysiology of mPFC circuits, leading to enhanced cognitive flexibility. All phenotypes only emerged in adulthood. Using viral Cre, we demonstrated that dcc organizes mPFC wiring specifically during adolescence. Variations in DCC may determine differential predisposition to mPFC disorders in humans. Indeed, DCC expression is elevated in brains of antidepressant-free subjects who committed suicide.
Figures
References
-
- Kalsbeek A, Voorn P, Buijs RM, Pool CW, Uylings HB. Development of the dopaminergic innervation in the prefrontal cortex of the rat. J Comp Neurol. 1988;269:58–72. - PubMed
-
- Benes FM, Taylor JB, Cunningham MC. Convergence and plasticity of monoaminergic systems in the medial prefrontal cortex during the postnatal period: implications for the development of psychopathology. Cereb Cortex. 2000;10:1014–1027. - PubMed
-
- Rosenberg DR, Lewis DA. Postnatal maturation of the dopaminergic innervation of monkey prefrontal and motor cortices: a tyrosine hydroxylase immunohistochemical analysis. J Comp Neurol. 1995;358:383–400. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
