

Horizontal gene transfer can rescue prokaryotes from Muller's ratchet: benefit of DNA from dead cells and population subdivision
- PMID: 24347631
- PMCID: PMC3931566
- DOI: 10.1534/g3.113.009845
Horizontal gene transfer can rescue prokaryotes from Muller's ratchet: benefit of DNA from dead cells and population subdivision
Abstract
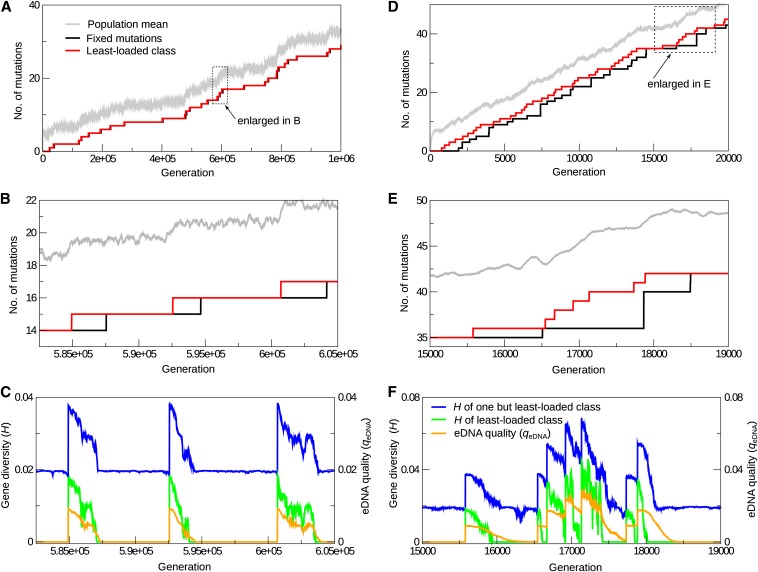
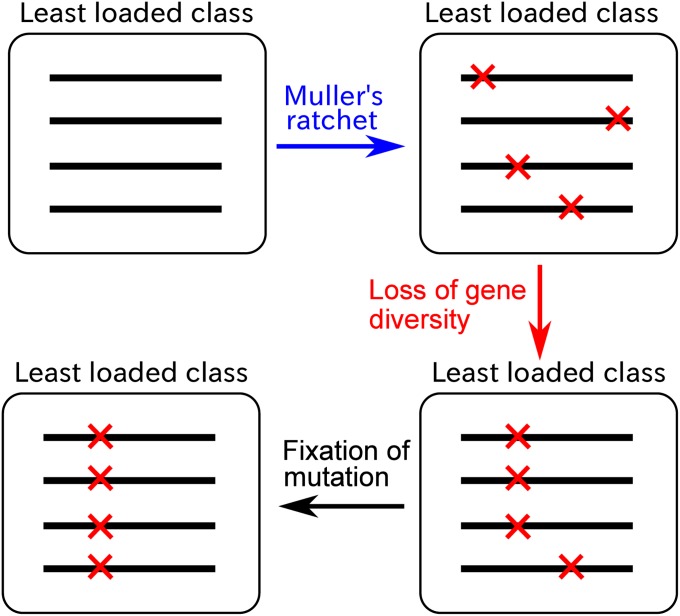
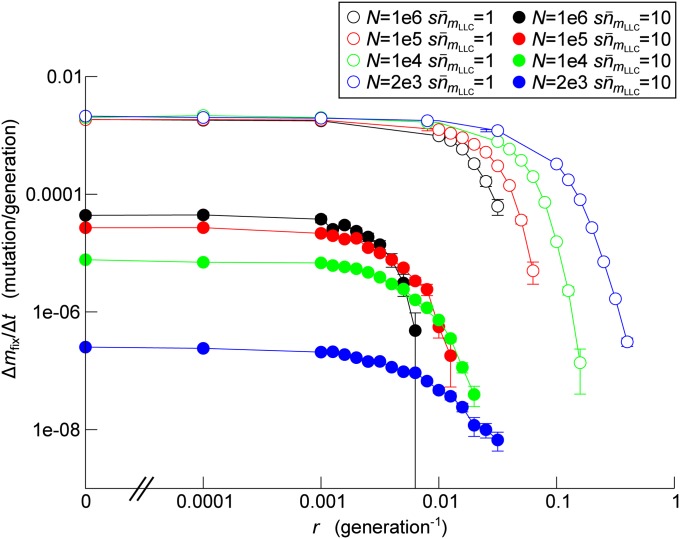
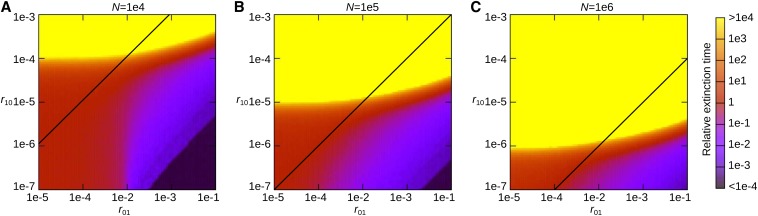
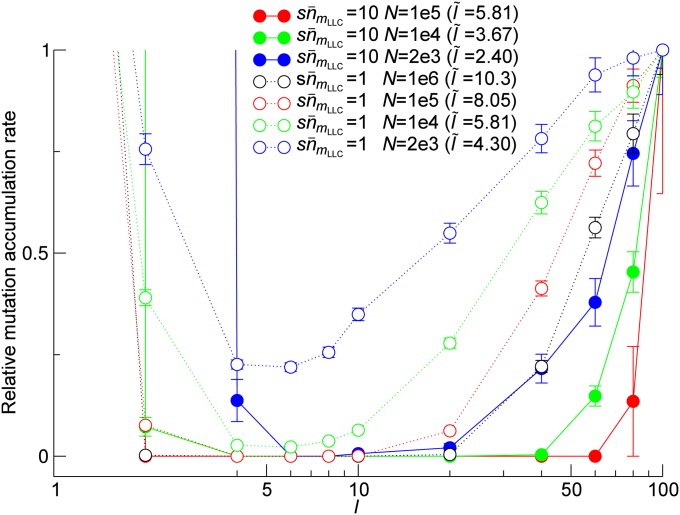
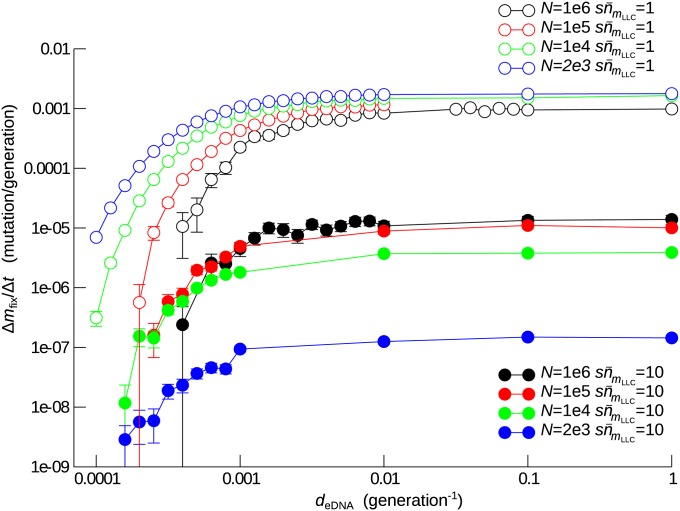
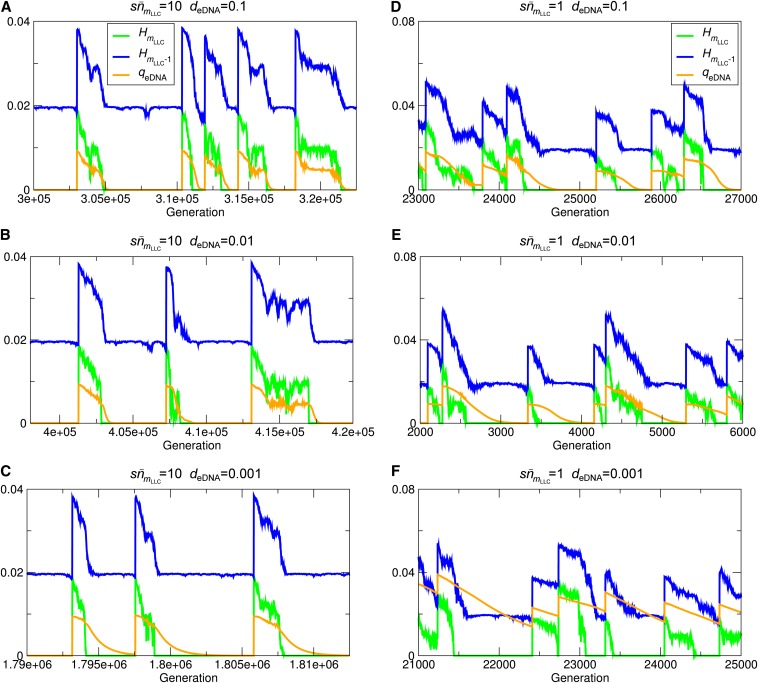
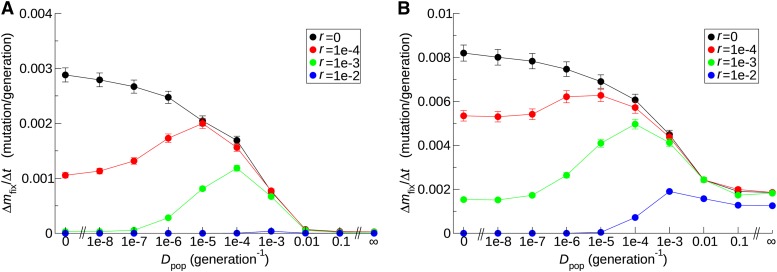
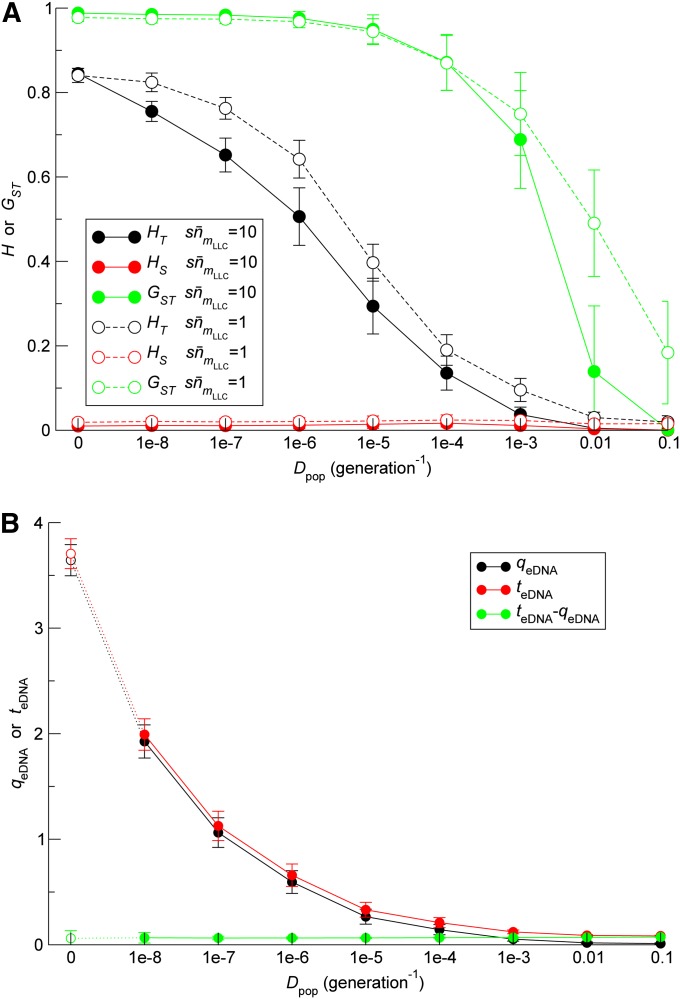
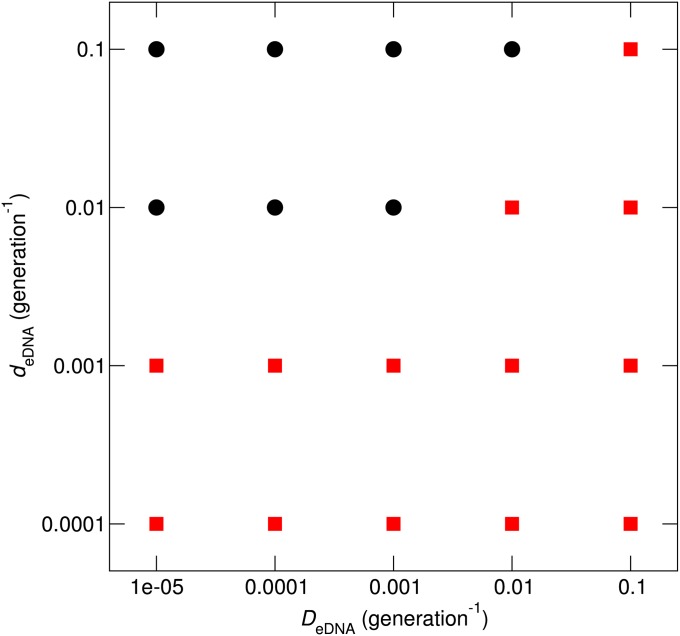
Horizontal gene transfer (HGT) is a major factor in the evolution of prokaryotes. An intriguing question is whether HGT is maintained during evolution of prokaryotes owing to its adaptive value or is a byproduct of selection driven by other factors such as consumption of extracellular DNA (eDNA) as a nutrient. One hypothesis posits that HGT can restore genes inactivated by mutations and thereby prevent stochastic, irreversible deterioration of genomes in finite populations known as Muller's ratchet. To examine this hypothesis, we developed a population genetic model of prokaryotes undergoing HGT via homologous recombination. Analysis of this model indicates that HGT can prevent the operation of Muller's ratchet even when the source of transferred genes is eDNA that comes from dead cells and on average carries more deleterious mutations than the DNA of recipient live cells. Moreover, if HGT is sufficiently frequent and eDNA diffusion sufficiently rapid, a subdivided population is shown to be more resistant to Muller's ratchet than an undivided population of an equal overall size. Thus, to maintain genomic information in the face of Muller's ratchet, it is more advantageous to partition individuals into multiple subpopulations and let them "cross-reference" each other's genetic information through HGT than to collect all individuals in one population and thereby maximize the efficacy of natural selection. Taken together, the results suggest that HGT could be an important condition for the long-term maintenance of genomic information in prokaryotes through the prevention of Muller's ratchet.
Keywords: competence; environmental DNA; evolution of transformation; soil bacteria; structured population.
Figures
References
-
- Aertsen A., Michiels C. W., 2004. Stress and how bacteria cope with death and survival. Crit. Rev. Microbiol. 30: 263–273 - PubMed
-
- Aravind L., Tatusov R. L., Wolf Y. I., Walker D. R., Koonin E. V., 1998. Evidence for massive gene exchange between archaeal and bacterial hyperthermophiles. Trends Genet. 14: 442–444 - PubMed
-
- Bell G., 1988. Recombination and the immortality of the germ line. J. Evol. Biol. 1: 67–82
-
- Bernstein H., Hopf F. A., Michod R. E., 1988. Is meiotic recombination an adaptation for repairing DNA, producing genetic variation, or both? pp. 139–160 in The Evolution of Sex, edited by Michod R. E., Levin B. R. Sinauer Associates, Sunderland, MA
-
- Blahna M. T., Zalewski C. A., Reuer J., Kahlmeter G., Foxman B., et al. , 2006. The role of horizontal gene transfer in the spread of trimethoprim-sulfamethoxazole resistance among uropathogenic Escherichia coli in Europe and Canada. J. Antimicrob. Chemother. 57: 666–672 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources