

Review
doi: 10.1155/2013/202358.
Epub 2013 Nov 19.
Close encounters of the third domain: the emerging genomic view of archaeal diversity and evolution
Affiliations
- PMID: 24348093
- PMCID: PMC3852633
- DOI: 10.1155/2013/202358
Item in Clipboard
Review
Close encounters of the third domain: the emerging genomic view of archaeal diversity and evolution
Archaea.
2013.
Abstract
The Archaea represent the so-called Third Domain of life, which has evolved in parallel with the Bacteria and which is implicated to have played a pivotal role in the emergence of the eukaryotic domain of life. Recent progress in genomic sequencing technologies and cultivation-independent methods has started to unearth a plethora of data of novel, uncultivated archaeal lineages. Here, we review how the availability of such genomic data has revealed several important insights into the diversity, ecological relevance, metabolic capacity, and the origin and evolution of the archaeal domain of life.
Figures
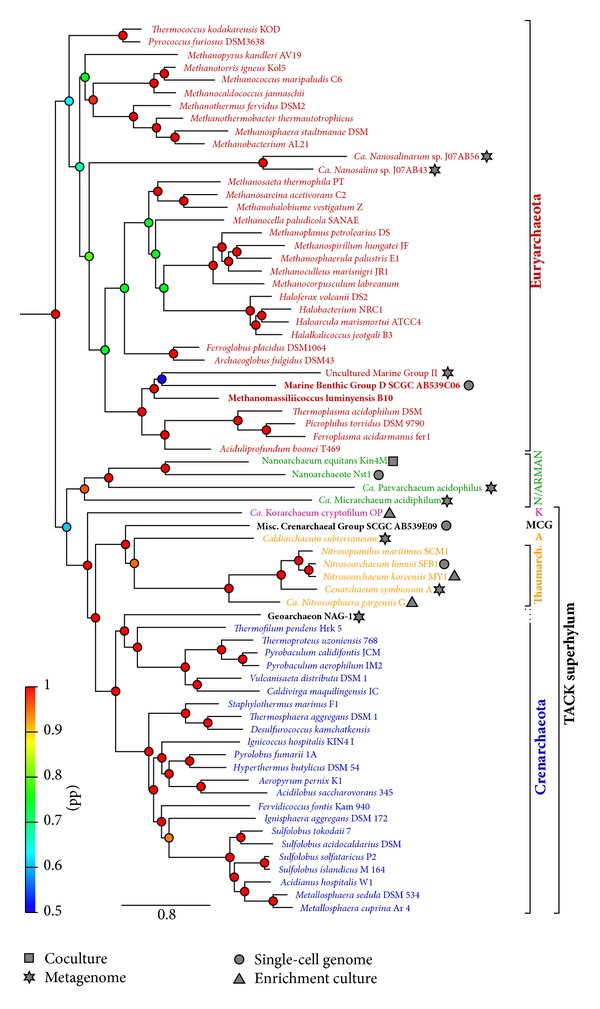
Bayesian phylogeny of 80 representative archaeal species. BLAST databases containing the proteome of 6 new archaeal genomes were retrieved from NCBI (in bold font on the tree): Methanomassiliicoccus luminyensis B10 (acc. no. CAJE01), MCG SCGC AB539E09 (acc. no. ALXK01), Marine Benthic Group D (MBGD) SCGC AB539N05, AB539C06, and AB540F20 (acc. no. ALXL01, AOSH01, and AOSI01, resp.). Protein sequence alignment from the 57 clusters in the discFilter 15 p dataset from [70] for which eukaryotes were removed were used as an input to psi-blast, with the six new proteomes as a database. Orthologs were retrieved as in [70]. For the three MBGD strains, one composite set of orthologs was constituted by using the most complete one (AB539C06) whenever possible and complementing with sequences from the other two if available. Orthologous genes selection, alignment, trimming, and concatenation were performed as in [70] resulting in a 15,069 amino-acid alignment. Four chains of Bayesian phylogenies were run with Phylobayes [71], under the CAT-Poisson model, running for approximately 10000 generations and discarding half as a burn-in. The tree was rooted with bacteria. Posterior probabilities (pp) are represented by colored dots on the nodes, with support values coloured according to the depicted heat-map colour scheme. The scale represents the number of substitutions per site. Species are colored according to the following: red, Euryarchaeota; green, Nanoarchaeota (N) and ARMAN; pink, Korarchaeota (K); black, Misc. Crenarchaeal Group (MCG); orange, Thaumarchaeota and Aigarchaeota (A); blue, Crenarchaeota. The DNA collection method, if different from pure culture, is indicated by a symbol next to the organism name: square, coculture; star, metagenome; circle, single-cell genome; triangle, enrichment culture.
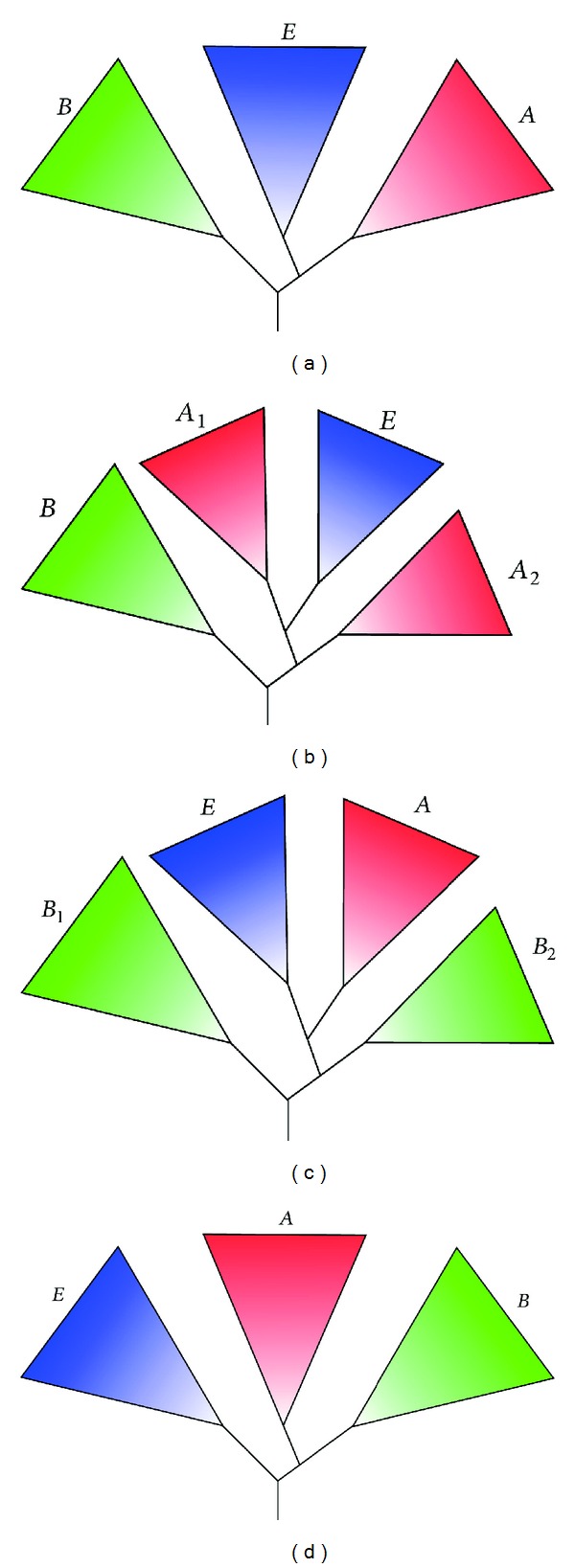
Overview of theories regarding the origin of the eukaryotic nuclear lineage. (a) The classical, Woesean three domains of life tree in which the nuclear lineage vertically evolved from the archaea-eukaryote common ancestor. (b) The fusion tree in which the nuclear lineage originated from the archaeal partner in the fusion. Depending on which fusion model, the archaeal parent's lineage (A
1) was either part of the euryarchaeota [SET [93], original syntrophy hypothesis [94], hydrogen hypothesis [95] or alternative syntrophy hypothesis [51]], the Crenarchaeota (eocyte hypothesis) [96], or the TACK superphylum (PhAT) [58]. “A
2” represents all archaea not directly affiliated with “A
1.” (c) The neomuran tree in which the eukaryotic and archaeal lineage (combined referred to as “neomurans”), evolved vertically from ancestor shared with actinobacteria (B
2) as a result of the loss of bacterial-type cell wall (the neomuran revolution). B
1 represents all bacteria not directly affiliated with B
2. (d) The eukaryote-early tree, which suggests that the last common universal ancestor was more eukaryote-like than prokaryote-like.
Similar articles
-
Genomic exploration of the diversity, ecology, and evolution of the archaeal domain of life.Science. 2017 Aug 11;357(6351):eaaf3883. doi: 10.1126/science.aaf3883. Science. 2017. PMID: 28798101 Review.
-
Genomic studies of uncultivated archaea.Nat Rev Microbiol. 2005 Jun;3(6):479-88. doi: 10.1038/nrmicro1159. Nat Rev Microbiol. 2005. PMID: 15931166 Review.
-
Exploring microbial dark matter to resolve the deep archaeal ancestry of eukaryotes.Philos Trans R Soc Lond B Biol Sci. 2015 Sep 26;370(1678):20140328. doi: 10.1098/rstb.2014.0328. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 26323759 Free PMC article.
-
Diversity, ecology and evolution of Archaea.Nat Microbiol. 2020 Jul;5(7):887-900. doi: 10.1038/s41564-020-0715-z. Epub 2020 May 4. Nat Microbiol. 2020. PMID: 32367054 Review.
-
Metagenomes from Coastal Marine Sediments Give Insights into the Ecological Role and Cellular Features of Loki- and Thorarchaeota.mBio. 2019 Sep 10;10(5):e02039-19. doi: 10.1128/mBio.02039-19. mBio. 2019. PMID: 31506313 Free PMC article.
Cited by
-
Fundamental issues related to the origin of melatonin and melatonin isomers during evolution: relation to their biological functions.Int J Mol Sci. 2014 Sep 9;15(9):15858-90. doi: 10.3390/ijms150915858. Int J Mol Sci. 2014. PMID: 25207599 Free PMC article. Review.
-
Astroglial vesicular network: evolutionary trends, physiology and pathophysiology.Acta Physiol (Oxf). 2018 Feb;222(2):10.1111/apha.12915. doi: 10.1111/apha.12915. Epub 2017 Aug 3. Acta Physiol (Oxf). 2018. PMID: 28665546 Free PMC article. Review.
-
Liposomes as vaccine delivery systems: a review of the recent advances.Ther Adv Vaccines. 2014 Nov;2(6):159-82. doi: 10.1177/2051013614541440. Ther Adv Vaccines. 2014. PMID: 25364509 Free PMC article. Review.
-
Nanotechnology Platform for Advancing Vaccine Development against the COVID-19 Virus.Diseases. 2023 Dec 10;11(4):177. doi: 10.3390/diseases11040177. Diseases. 2023. PMID: 38131983 Free PMC article. Review.
-
Microorganisms-A Journal and a Unifying Concept for the Science of Microbiology.Microorganisms. 2014 Dec 12;2(4):140-6. doi: 10.3390/microorganisms2040140. Microorganisms. 2014. PMID: 27682235 Free PMC article.
References
-
- Woese CR. The birth of the Archaea: a personal retrospective. In: Garrett RA, Klenk H-P, editors. Archaea. Blackwell Publishing; 2007.
-
- Kandler O, Hippe H. Lack of peptidoglycan in the cell walls of Methanosarcina barkeri. Archives of Microbiology. 1977;113(1-2):57–60. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
