

Inferring developmental stage composition from gene expression in human malaria
- PMID: 24348235
- PMCID: PMC3861035
- DOI: 10.1371/journal.pcbi.1003392
Inferring developmental stage composition from gene expression in human malaria
Abstract
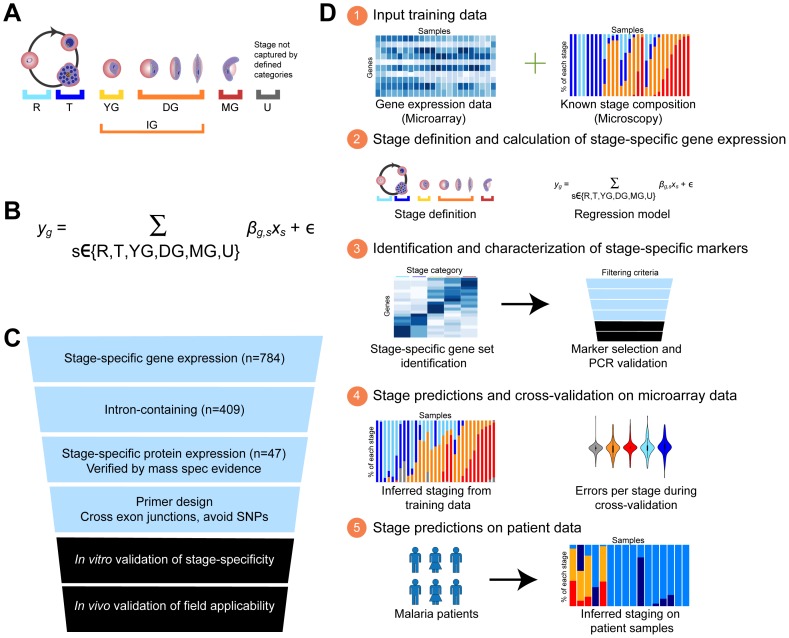
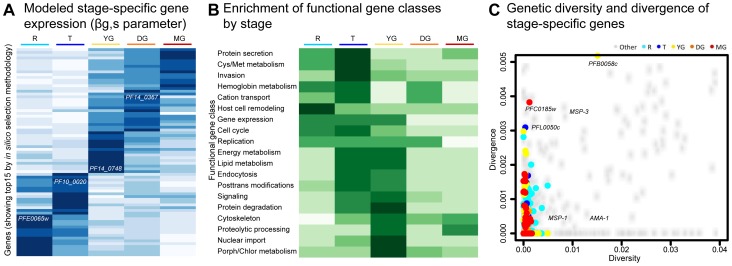
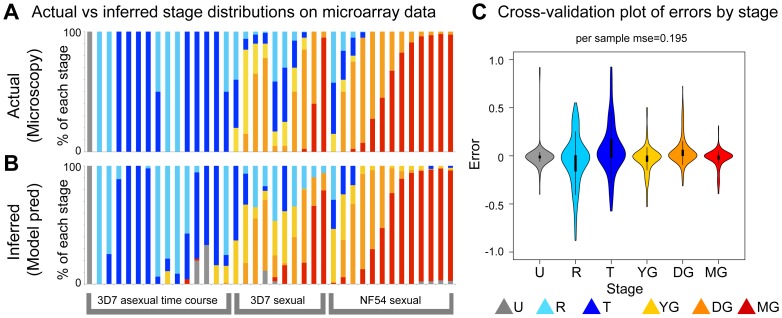
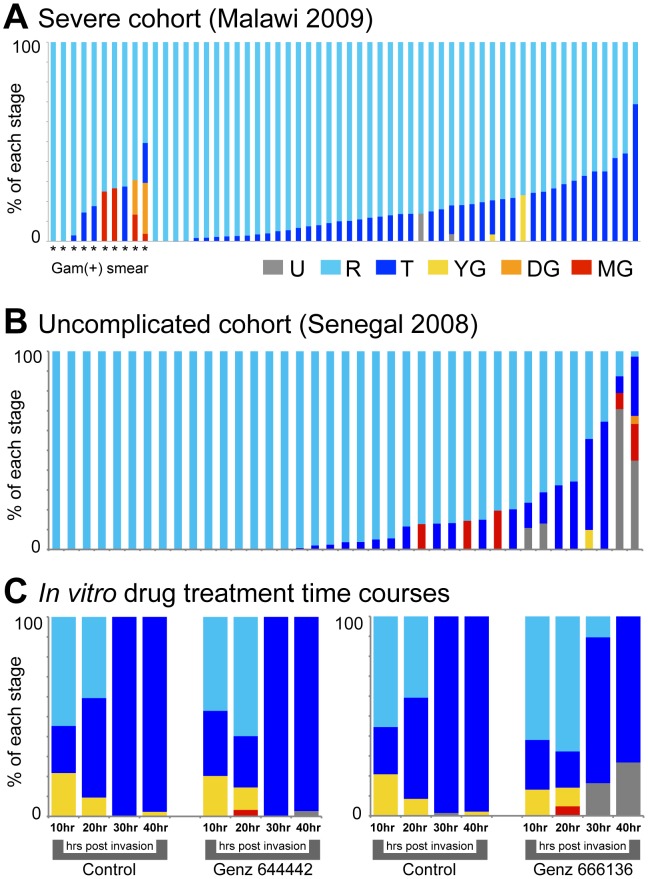
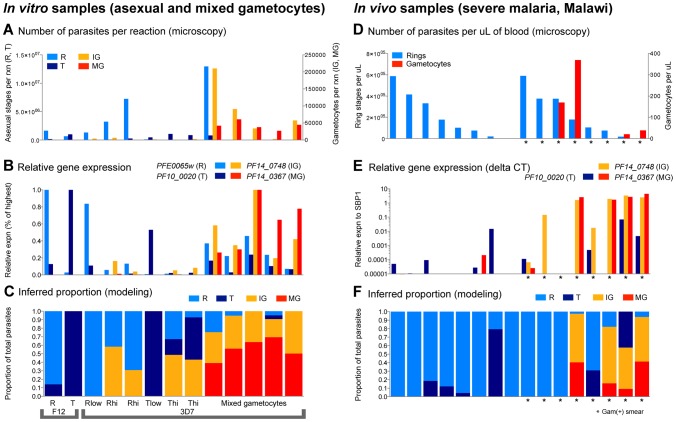
In the current era of malaria eradication, reducing transmission is critical. Assessment of transmissibility requires tools that can accurately identify the various developmental stages of the malaria parasite, particularly those required for transmission (sexual stages). Here, we present a method for estimating relative amounts of Plasmodium falciparum asexual and sexual stages from gene expression measurements. These are modeled using constrained linear regression to characterize stage-specific expression profiles within mixed-stage populations. The resulting profiles were analyzed functionally by gene set enrichment analysis (GSEA), confirming differentially active pathways such as increased mitochondrial activity and lipid metabolism during sexual development. We validated model predictions both from microarrays and from quantitative RT-PCR (qRT-PCR) measurements, based on the expression of a small set of key transcriptional markers. This sufficient marker set was identified by backward selection from the whole genome as available from expression arrays, targeting one sentinel marker per stage. The model as learned can be applied to any new microarray or qRT-PCR transcriptional measurement. We illustrate its use in vitro in inferring changes in stage distribution following stress and drug treatment and in vivo in identifying immature and mature sexual stage carriers within patient cohorts. We believe this approach will be a valuable resource for staging lab and field samples alike and will have wide applicability in epidemiological studies of malaria transmission.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Alano P (2007) Plasmodium falciparum gametocytes: still many secrets of a hidden life. Mol Microbiol 66: 291–302. - PubMed
-
- Young JA, Fivelman QL, Blair PL, de la Vega P, Le Roch KG, et al. (2005) The Plasmodium falciparum sexual development transcriptome: a microarray analysis using ontology-based pattern identification. Mol Biochem Parasitol 143: 67–79. - PubMed
-
- Le Roch KG, Zhou Y, Blair PL, Grainger M, Moch JK, et al. (2003) Discovery of gene function by expression profiling of the malaria parasite life cycle. Science 301: 1503–1508. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
