

A global in vivo Drosophila RNAi screen identifies a key role of ceramide phosphoethanolamine for glial ensheathment of axons
- PMID: 24348263
- PMCID: PMC3861124
- DOI: 10.1371/journal.pgen.1003980
A global in vivo Drosophila RNAi screen identifies a key role of ceramide phosphoethanolamine for glial ensheathment of axons
Abstract
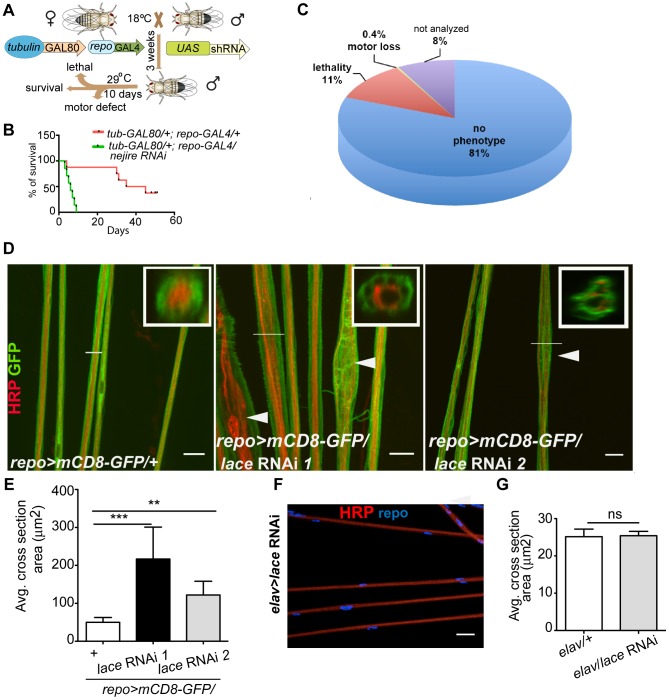
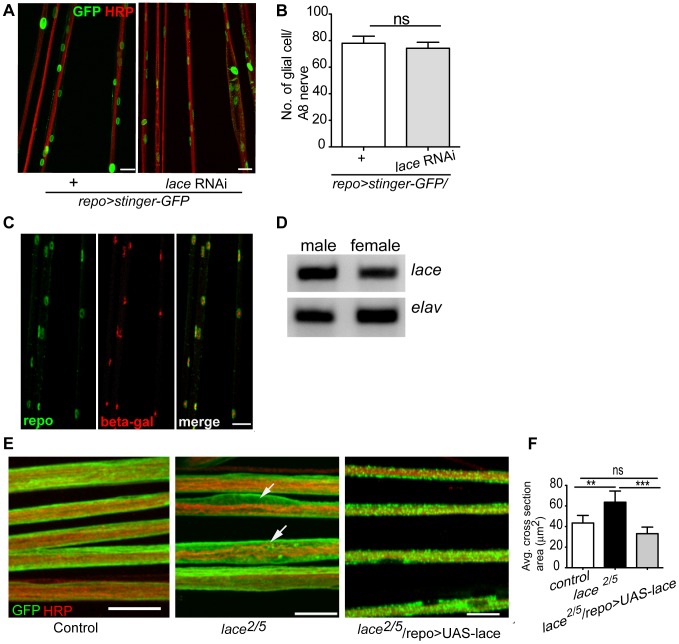
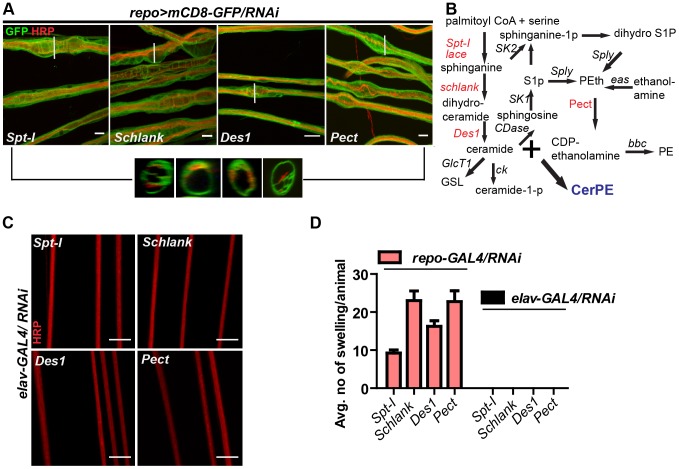
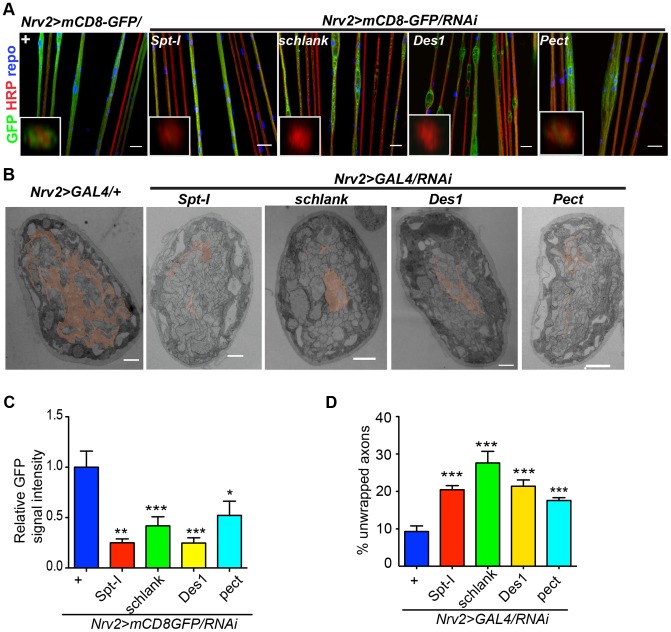
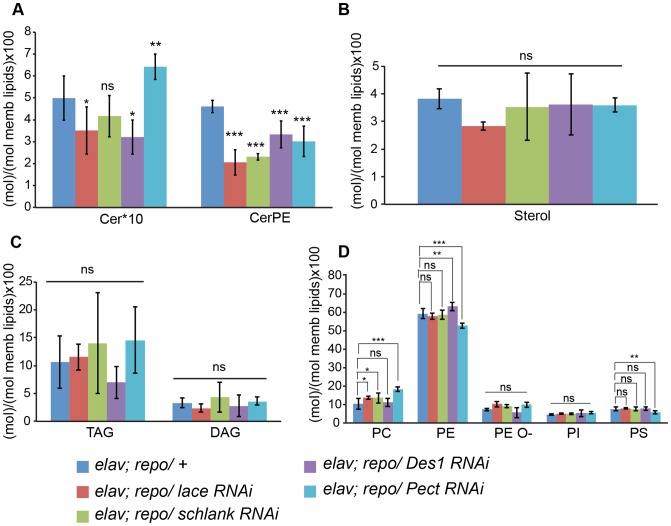
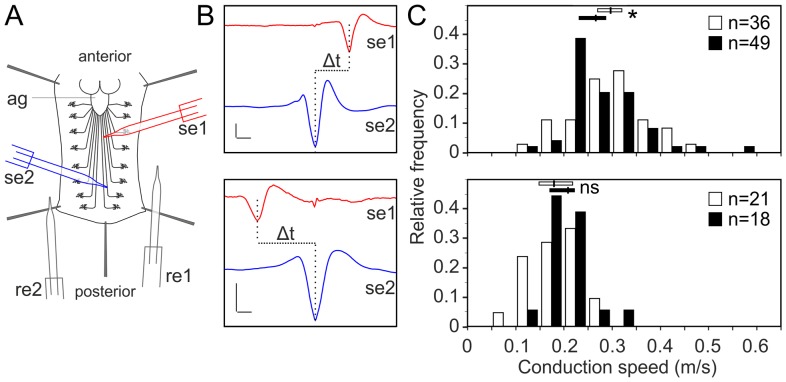
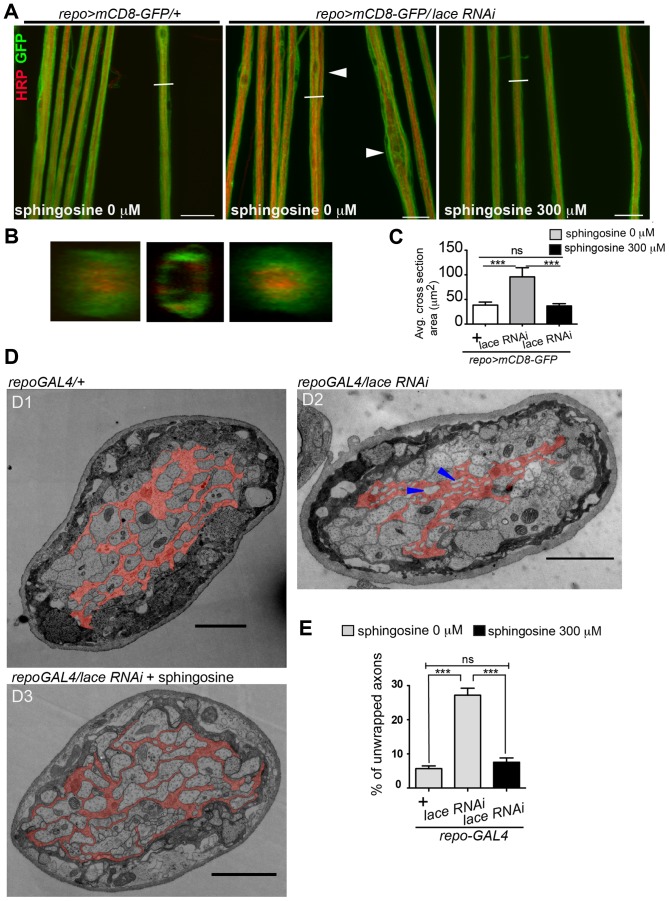
Glia are of vital importance for all complex nervous system. One of the many functions of glia is to insulate and provide trophic and metabolic support to axons. Here, using glial-specific RNAi knockdown in Drosophila, we silenced 6930 conserved genes in adult flies to identify essential genes and pathways. Among our screening hits, metabolic processes were highly represented, and genes involved in carbohydrate and lipid metabolic pathways appeared to be essential in glia. One critical pathway identified was de novo ceramide synthesis. Glial knockdown of lace, a subunit of the serine palmitoyltransferase associated with hereditary sensory and autonomic neuropathies in humans, resulted in ensheathment defects of peripheral nerves in Drosophila. A genetic dissection study combined with shotgun high-resolution mass spectrometry of lipids showed that levels of ceramide phosphoethanolamine are crucial for axonal ensheathment by glia. A detailed morphological and functional analysis demonstrated that the depletion of ceramide phosphoethanolamine resulted in axonal defasciculation, slowed spike propagation, and failure of wrapping glia to enwrap peripheral axons. Supplementing sphingosine into the diet rescued the neuropathy in flies. Thus, our RNAi study in Drosophila identifies a key role of ceramide phosphoethanolamine in wrapping of axons by glia.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Peripheral axonal ensheathment is regulated by RalA GTPase and the exocyst complex.Development. 2020 Feb 4;147(3):dev174540. doi: 10.1242/dev.174540. Development. 2020. PMID: 31969325
-
Axonal ensheathment and septate junction formation in the peripheral nervous system of Drosophila.J Neurosci. 2006 Mar 22;26(12):3319-29. doi: 10.1523/JNEUROSCI.5383-05.2006. J Neurosci. 2006. PMID: 16554482 Free PMC article.
-
Borderless regulates glial extension and axon ensheathment.Dev Biol. 2016 Jun 15;414(2):170-80. doi: 10.1016/j.ydbio.2016.04.020. Epub 2016 Apr 27. Dev Biol. 2016. PMID: 27131624
-
Wrapping axons in mammals and Drosophila: Different lipids, same principle.Biochimie. 2020 Nov;178:39-48. doi: 10.1016/j.biochi.2020.08.002. Epub 2020 Aug 13. Biochimie. 2020. PMID: 32800899 Review.
-
Glial ensheathment of peripheral axons in Drosophila.J Neurosci Res. 2008 May 1;86(6):1189-98. doi: 10.1002/jnr.21574. J Neurosci Res. 2008. PMID: 18041093 Free PMC article. Review.
Cited by
-
Loss of Frataxin induces iron toxicity, sphingolipid synthesis, and Pdk1/Mef2 activation, leading to neurodegeneration.Elife. 2016 Jun 25;5:e16043. doi: 10.7554/eLife.16043. Elife. 2016. PMID: 27343351 Free PMC article.
-
Variable Substrate Preference among Phospholipase D Toxins from Sicariid Spiders.J Biol Chem. 2015 Apr 24;290(17):10994-1007. doi: 10.1074/jbc.M115.636951. Epub 2015 Mar 9. J Biol Chem. 2015. PMID: 25752604 Free PMC article.
-
Discoidin domain receptor regulates ensheathment, survival and caliber of peripheral axons.Development. 2022 Dec 1;149(23):dev200636. doi: 10.1242/dev.200636. Epub 2022 Dec 13. Development. 2022. PMID: 36355066 Free PMC article.
-
Peroxisomes are required for lipid metabolism and muscle function in Drosophila melanogaster.PLoS One. 2014 Jun 19;9(6):e100213. doi: 10.1371/journal.pone.0100213. eCollection 2014. PLoS One. 2014. PMID: 24945818 Free PMC article.
-
The Little Fly that Could: Wizardry and Artistry of Drosophila Genomics.Genes (Basel). 2014 May 13;5(2):385-414. doi: 10.3390/genes5020385. Genes (Basel). 2014. PMID: 24827974 Free PMC article.
References
-
- Freeman MR, Doherty J (2006) Glial cell biology in Drosophila and vertebrates. Trends Neurosci 29: 82–90. - PubMed
-
- Barres BA (2008) The mystery and magic of glia: a perspective on their roles in health and disease. Neuron 60: 430–440. - PubMed
-
- Sherman DL, Brophy PJ (2005) Mechanisms of axon ensheathment and myelin growth. Nat Rev Neurosci 6: 683–690. - PubMed
-
- Mirsky R, Jessen KR (1996) Schwann cell development, differentiation and myelination. Curr Opin Neurobiol 6: 89–96. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
