

Ethylene promotes hypocotyl growth and HY5 degradation by enhancing the movement of COP1 to the nucleus in the light
- PMID: 24348273
- PMCID: PMC3861121
- DOI: 10.1371/journal.pgen.1004025
Ethylene promotes hypocotyl growth and HY5 degradation by enhancing the movement of COP1 to the nucleus in the light
Abstract
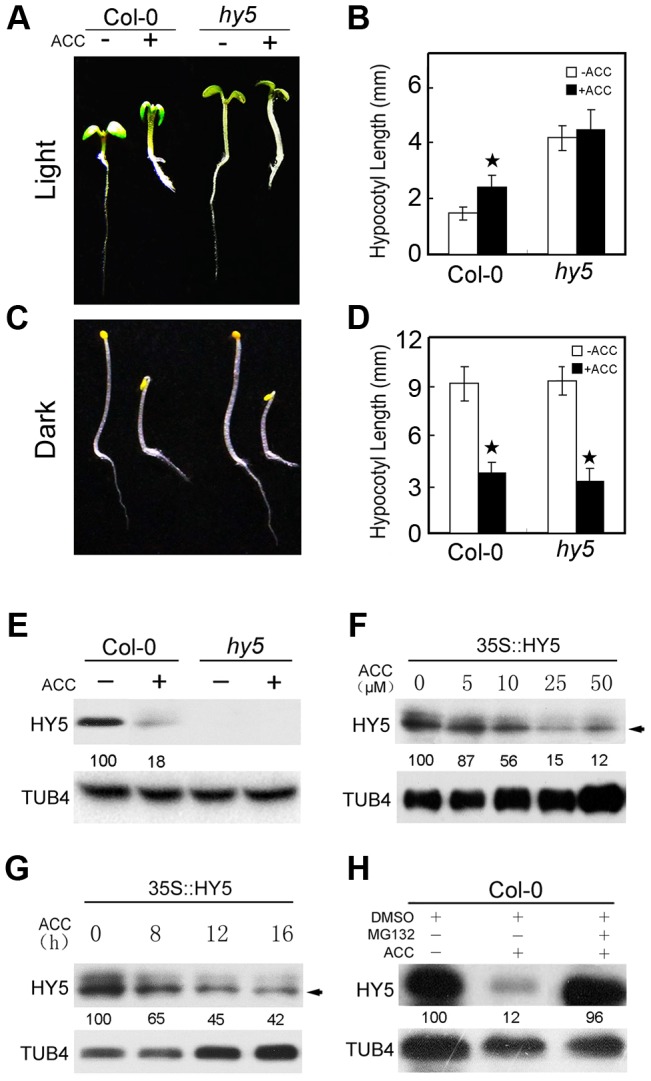
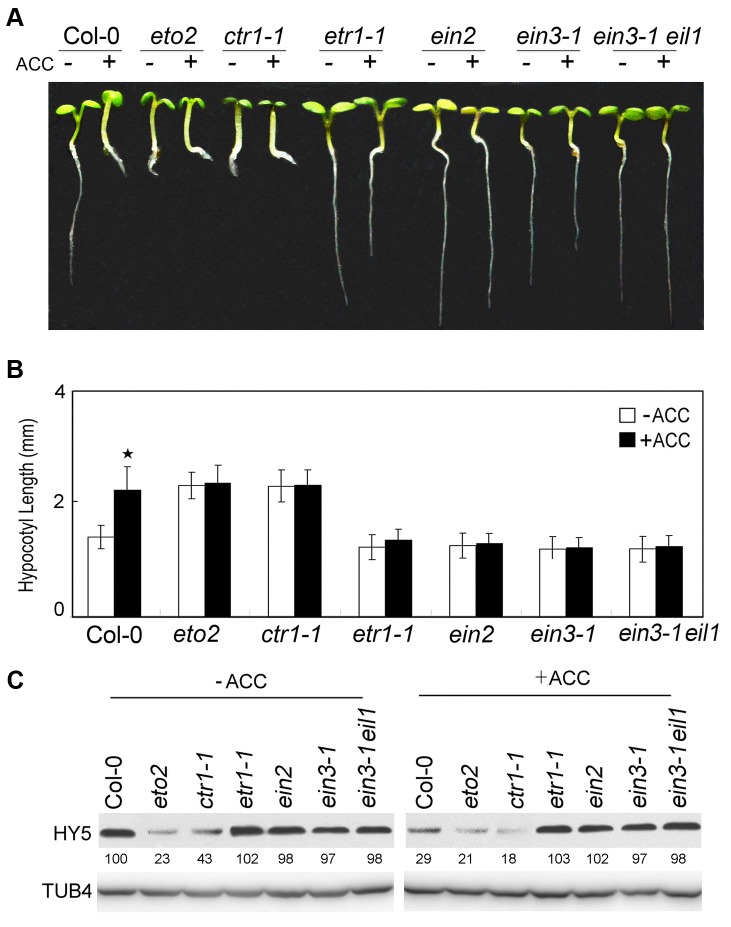
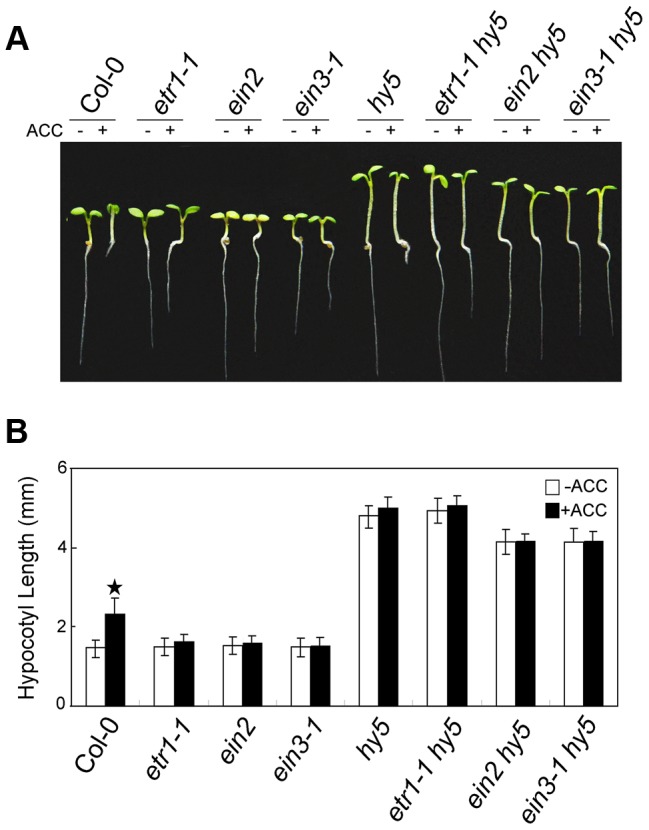
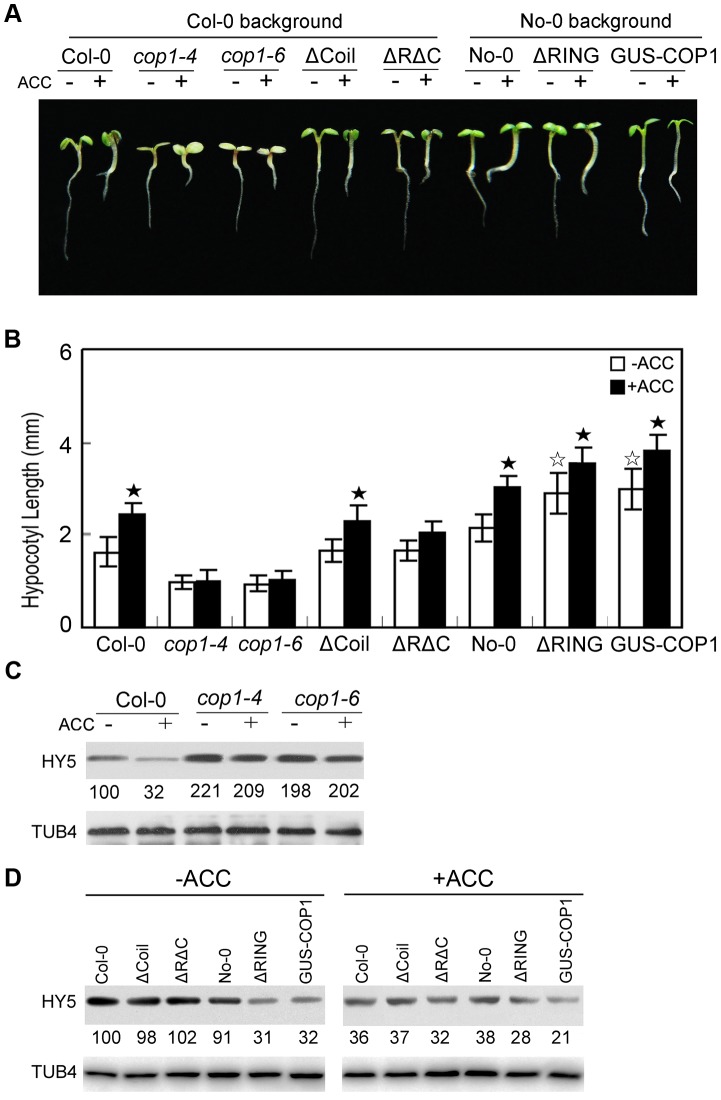
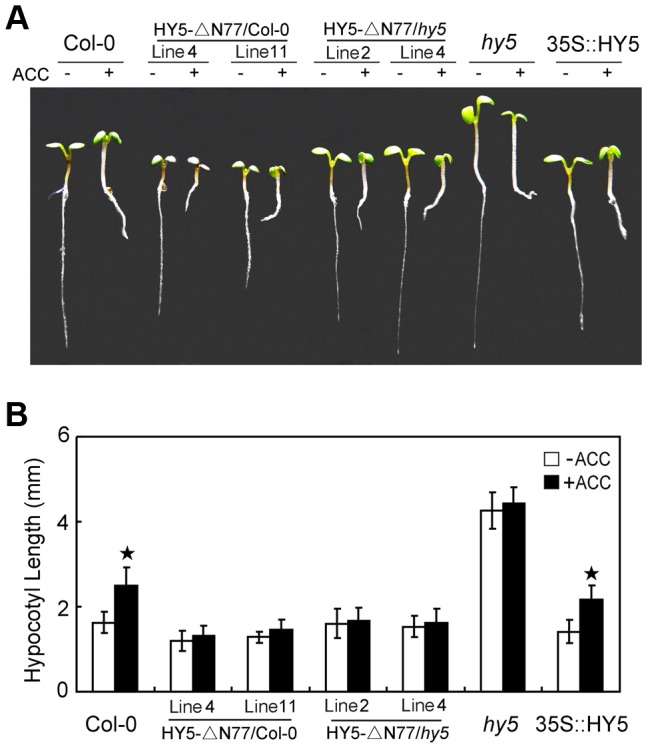
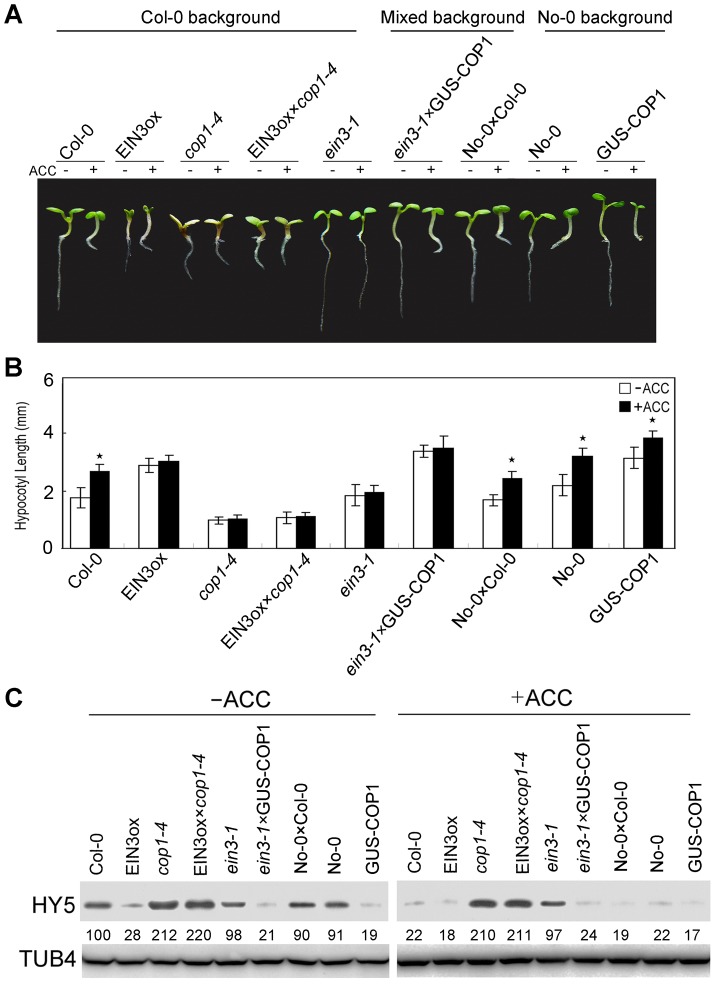
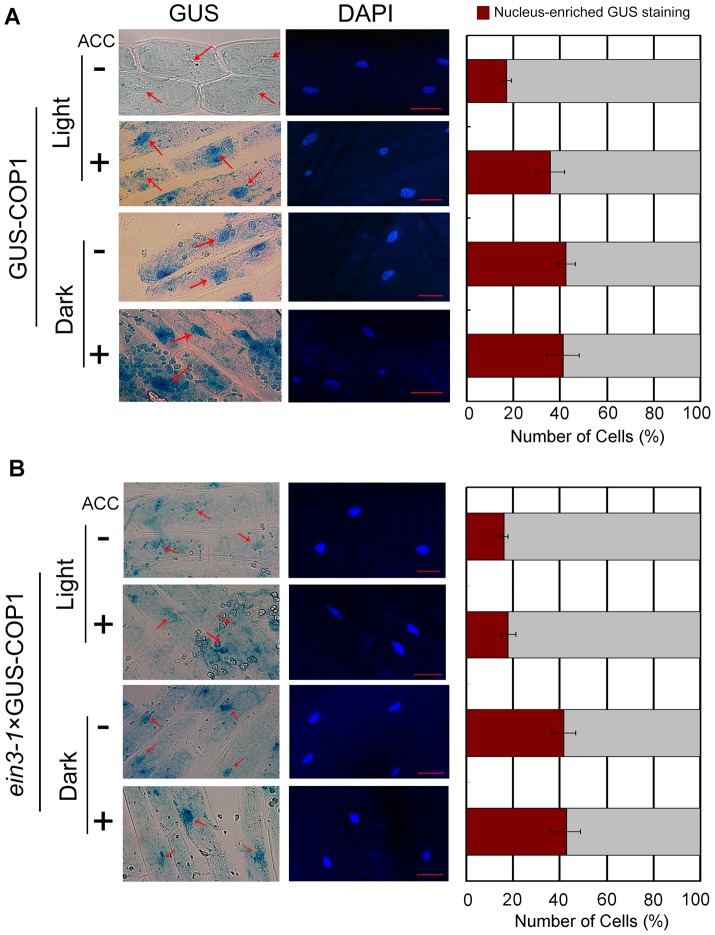
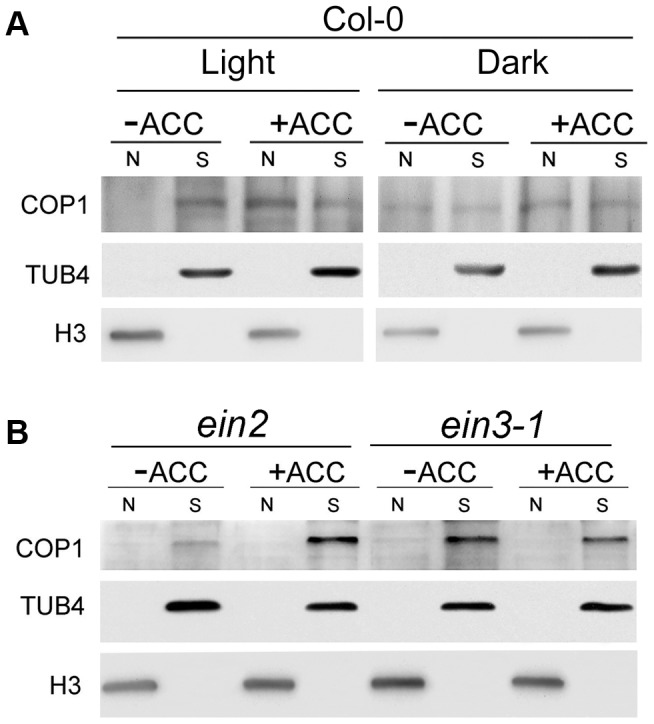
In the dark, etiolated seedlings display a long hypocotyl, the growth of which is rapidly inhibited when the seedlings are exposed to light. In contrast, the phytohormone ethylene prevents hypocotyl elongation in the dark but enhances its growth in the light. However, the mechanism by which light and ethylene signalling oppositely affect this process at the protein level is unclear. Here, we report that ethylene enhances the movement of constitutive photomorphogenesis 1 (COP1) to the nucleus where it mediates the degradation of long hypocotyl 5 (HY5), contributing to hypocotyl growth in the light. Our results indicate that HY5 is required for ethylene-promoted hypocotyl growth in the light, but not in the dark. Using genetic and biochemical analyses, we found that HY5 functions downstream of ethylene insensitive 3 (EIN3) for ethylene-promoted hypocotyl growth. Furthermore, the upstream regulation of HY5 stability by ethylene is COP1-dependent, and COP1 is genetically located downstream of EIN3, indicating that the COP1-HY5 complex integrates light and ethylene signalling downstream of EIN3. Importantly, the ethylene precursor 1-aminocyclopropane-1-carboxylate (ACC) enriched the nuclear localisation of COP1; however, this effect was dependent on EIN3 only in the presence of light, strongly suggesting that ethylene promotes the effects of light on the movement of COP1 from the cytoplasm to the nucleus. Thus, our investigation demonstrates that the COP1-HY5 complex is a novel integrator that plays an essential role in ethylene-promoted hypocotyl growth in the light.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Involvement of COP1 in ethylene- and light-regulated hypocotyl elongation.Planta. 2012 Dec;236(6):1791-802. doi: 10.1007/s00425-012-1730-y. Epub 2012 Aug 14. Planta. 2012. PMID: 22890836
-
Short Hypocotyl in White Light1 Interacts with Elongated Hypocotyl5 (HY5) and Constitutive Photomorphogenic1 (COP1) and Promotes COP1-Mediated Degradation of HY5 during Arabidopsis Seedling Development.Plant Physiol. 2015 Dec;169(4):2922-34. doi: 10.1104/pp.15.01184. Epub 2015 Oct 16. Plant Physiol. 2015. PMID: 26474641 Free PMC article.
-
COP1-mediated degradation of BBX22/LZF1 optimizes seedling development in Arabidopsis.Plant Physiol. 2011 May;156(1):228-39. doi: 10.1104/pp.111.175042. Epub 2011 Mar 22. Plant Physiol. 2011. PMID: 21427283 Free PMC article.
-
COP1 and BBXs-HY5-mediated light signal transduction in plants.New Phytol. 2020 Dec;228(6):1748-1753. doi: 10.1111/nph.16296. Epub 2019 Dec 7. New Phytol. 2020. PMID: 31664720 Review.
-
Seedlings Transduce the Depth and Mechanical Pressure of Covering Soil Using COP1 and Ethylene to Regulate EBF1/EBF2 for Soil Emergence.Curr Biol. 2016 Jan 25;26(2):139-149. doi: 10.1016/j.cub.2015.11.053. Epub 2015 Dec 31. Curr Biol. 2016. PMID: 26748855 Free PMC article. Review.
Cited by
-
DNA methylation and transcriptomic changes in response to different lights and stresses in 7B-1 male-sterile tomato.PLoS One. 2015 Apr 7;10(4):e0121864. doi: 10.1371/journal.pone.0121864. eCollection 2015. PLoS One. 2015. PMID: 25849771 Free PMC article.
-
Phytochrome and Phytohormones: Working in Tandem for Plant Growth and Development.Front Plant Sci. 2018 Jul 27;9:1037. doi: 10.3389/fpls.2018.01037. eCollection 2018. Front Plant Sci. 2018. PMID: 30100912 Free PMC article. Review.
-
CRISPR/Cas9-Mediated HY5 Gene Editing Reduces Growth Inhibition in Chinese Cabbage (Brassica rapa) under ER Stress.Int J Mol Sci. 2023 Aug 23;24(17):13105. doi: 10.3390/ijms241713105. Int J Mol Sci. 2023. PMID: 37685921 Free PMC article.
-
Light and Ethylene Coordinately Regulate the Phosphate Starvation Response through Transcriptional Regulation of PHOSPHATE STARVATION RESPONSE1.Plant Cell. 2017 Sep;29(9):2269-2284. doi: 10.1105/tpc.17.00268. Epub 2017 Aug 25. Plant Cell. 2017. PMID: 28842534 Free PMC article.
-
Light Modulates Ethylene Synthesis, Signaling, and Downstream Transcriptional Networks to Control Plant Development.Front Plant Sci. 2019 Sep 12;10:1094. doi: 10.3389/fpls.2019.01094. eCollection 2019. Front Plant Sci. 2019. PMID: 31572414 Free PMC article. Review.
References
-
- Guo H, Ecker JR (2004) The ethylene signaling pathway: new insights. Curr Opin Plant Biol 7: 40–49. - PubMed
-
- Stepanova AN, Alonso JM (2009) Ethylene signaling and response: where different regulatory modules meet. Curr Opin Plant Biol 12: 548–555. - PubMed
-
- Hua J, Meyerowitz EM (1998) Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana . Cell 94: 261–271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
