

MicroRNA156: a potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena
- PMID: 24351688
- PMCID: PMC3912076
- DOI: 10.1104/pp.113.230714
MicroRNA156: a potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena
Abstract
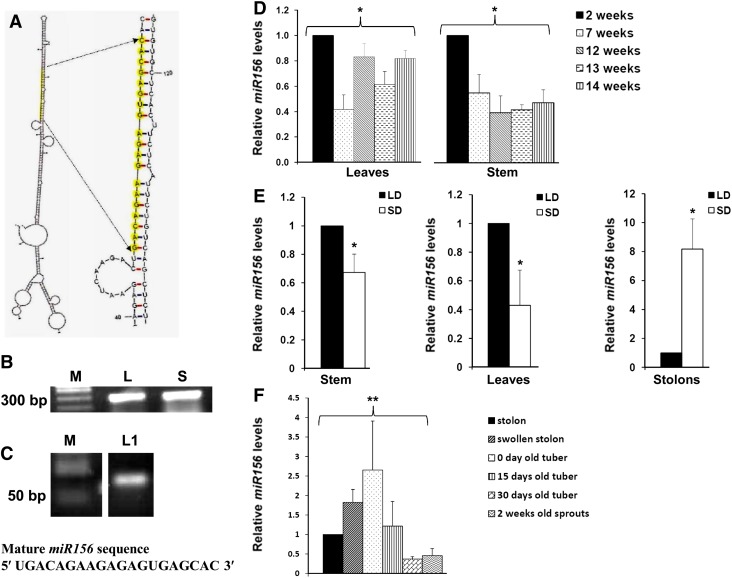
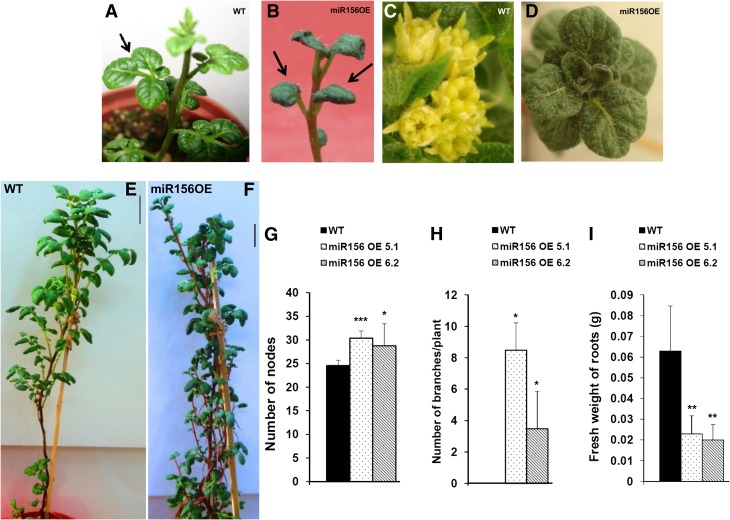
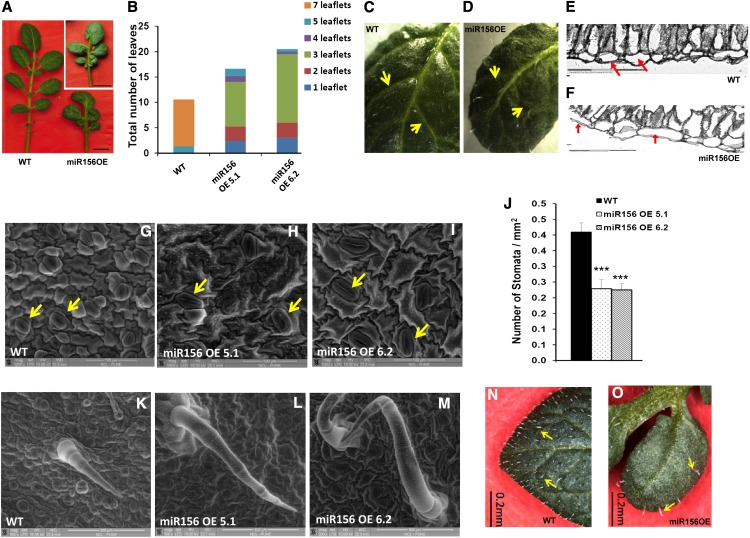
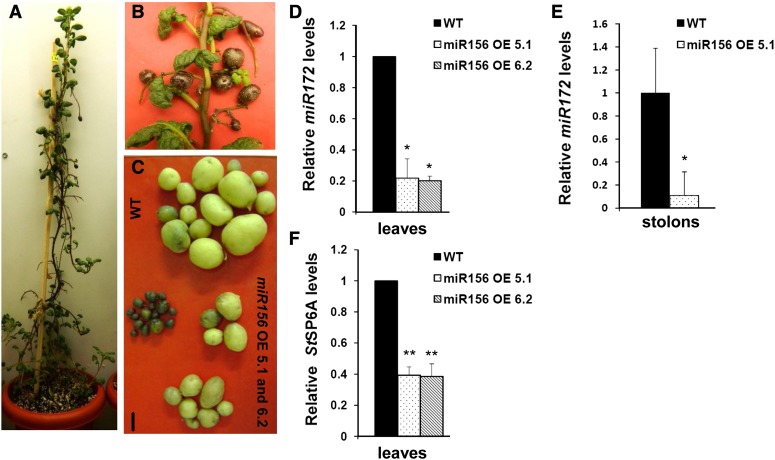
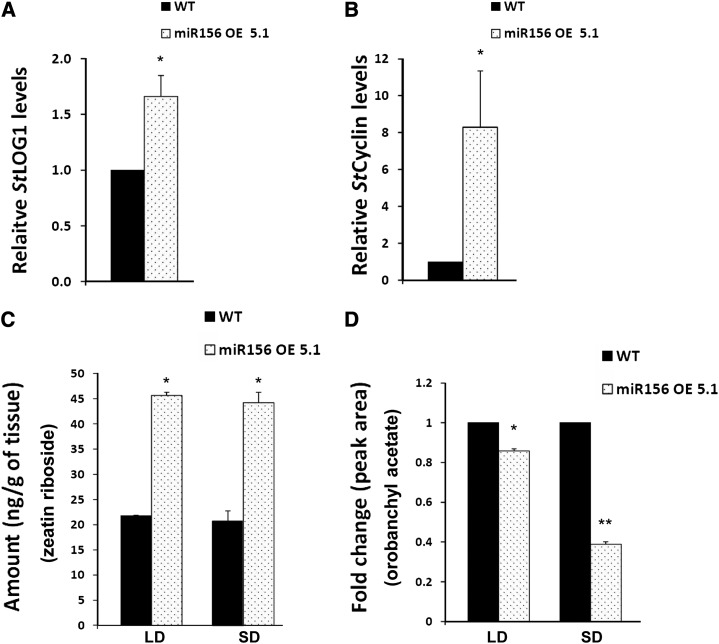
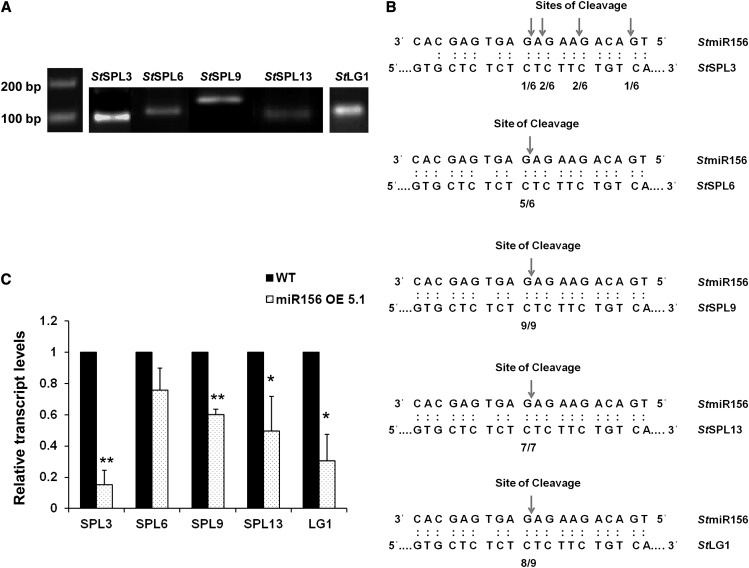
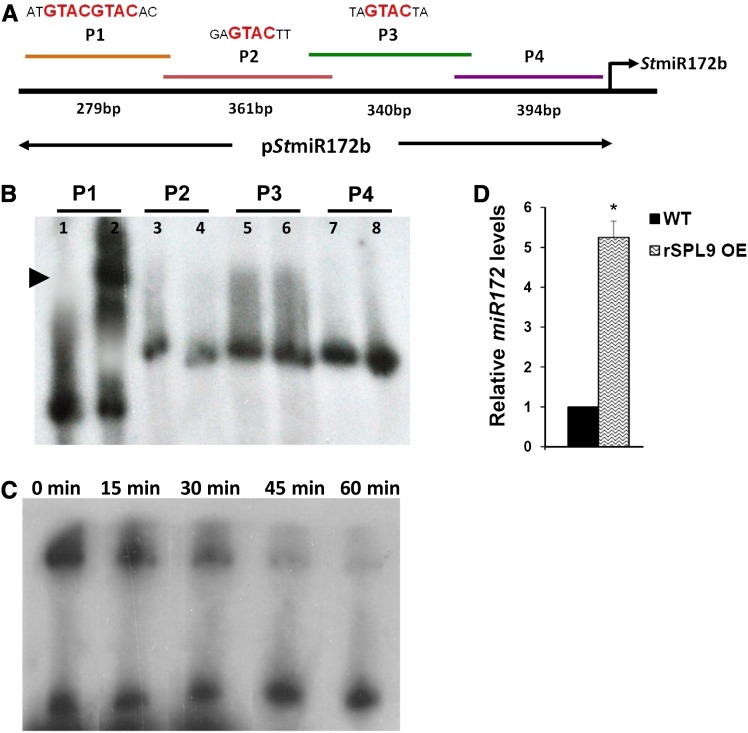
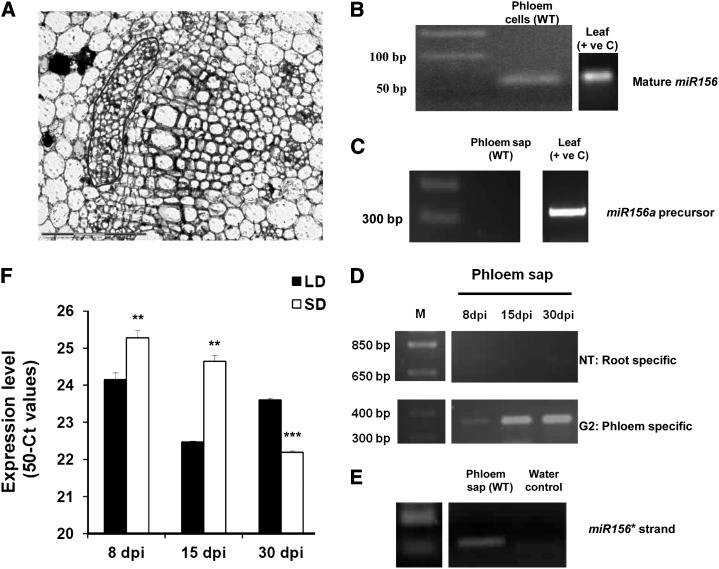
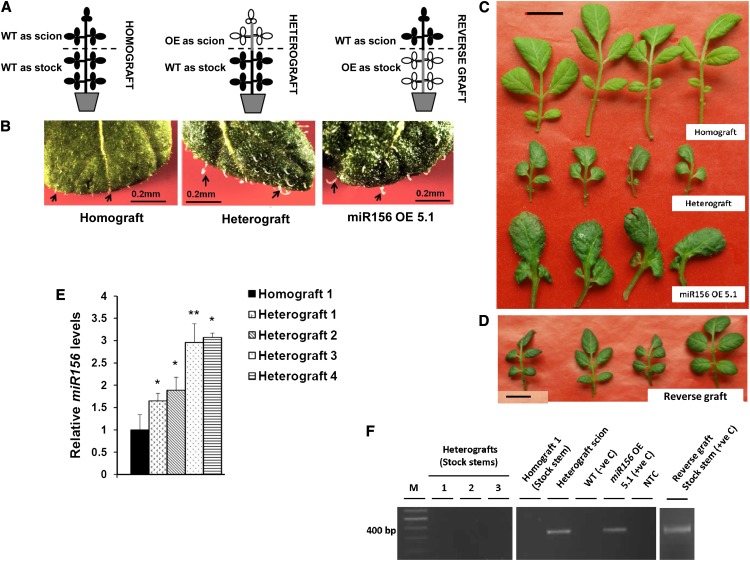
MicroRNA156 (miR156) functions in maintaining the juvenile phase in plants. However, the mobility of this microRNA has not been demonstrated. So far, only three microRNAs, miR399, miR395, and miR172, have been shown to be mobile. We demonstrate here that miR156 is a potential graft-transmissible signal that affects plant architecture and tuberization in potato (Solanum tuberosum). Under tuber-noninductive (long-day) conditions, miR156 shows higher abundance in leaves and stems, whereas an increase in abundance of miR156 has been observed in stolons under tuber-inductive (short-day) conditions, indicative of a photoperiodic control. Detection of miR156 in phloem cells of wild-type plants and mobility assays in heterografts suggest that miR156 is a graft-transmissible signal. This movement was correlated with changes in leaf morphology and longer trichomes in leaves. Overexpression of miR156 in potato caused a drastic phenotype resulting in altered plant architecture and reduced tuber yield. miR156 overexpression plants also exhibited altered levels of cytokinin and strigolactone along with increased levels of LONELY GUY1 and StCyclin D3.1 transcripts as compared with wild-type plants. RNA ligase-mediated rapid amplification of complementary DNA ends analysis validated SQUAMOSA PROMOTER BINDING-LIKE3 (StSPL3), StSPL6, StSPL9, StSPL13, and StLIGULELESS1 as targets of miR156. Gel-shift assays indicate the regulation of miR172 by miR156 through StSPL9. miR156-resistant SPL9 overexpression lines exhibited increased miR172 levels under a short-day photoperiod, supporting miR172 regulation via the miR156-SPL9 module. Overall, our results strongly suggest that miR156 is a phloem-mobile signal regulating potato development.
Figures
References
-
- Atkins CA, Smith PMC, Rodriguez-Medina C. (2011) Macromolecules in phloem exudates: a review. Protoplasma 248: 165–172 - PubMed
-
- Axtell MJ, Bowman JL. (2008) Evolution of plant microRNAs and their targets. Trends Plant Sci 13: 343–349 - PubMed
-
- Banerjee AK, Prat S, Hannapel DJ. (2006b) Efficient production of transgenic potato (S. tuberosum L. ssp. andigena) plants via Agrobacterium tumefaciens-mediated transformation. Plant Sci 170: 732–738
-
- Birkenbihl RP, Jach G, Saedler H, Huijser P. (2005) Functional dissection of the plant-specific SBP-domain: overlap of the DNA-binding and nuclear localization domains. J Mol Biol 352: 585–596 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
