

An ITAM in a nonenveloped virus regulates activation of NF-κB, induction of beta interferon, and viral spread
- PMID: 24352448
- PMCID: PMC3958081
- DOI: 10.1128/JVI.02573-13
An ITAM in a nonenveloped virus regulates activation of NF-κB, induction of beta interferon, and viral spread
Abstract
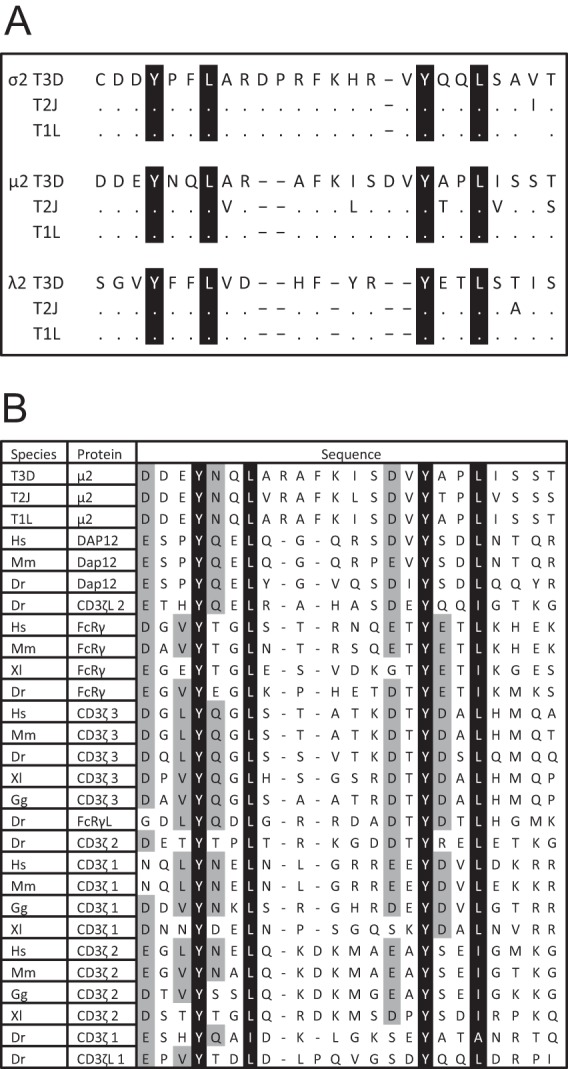
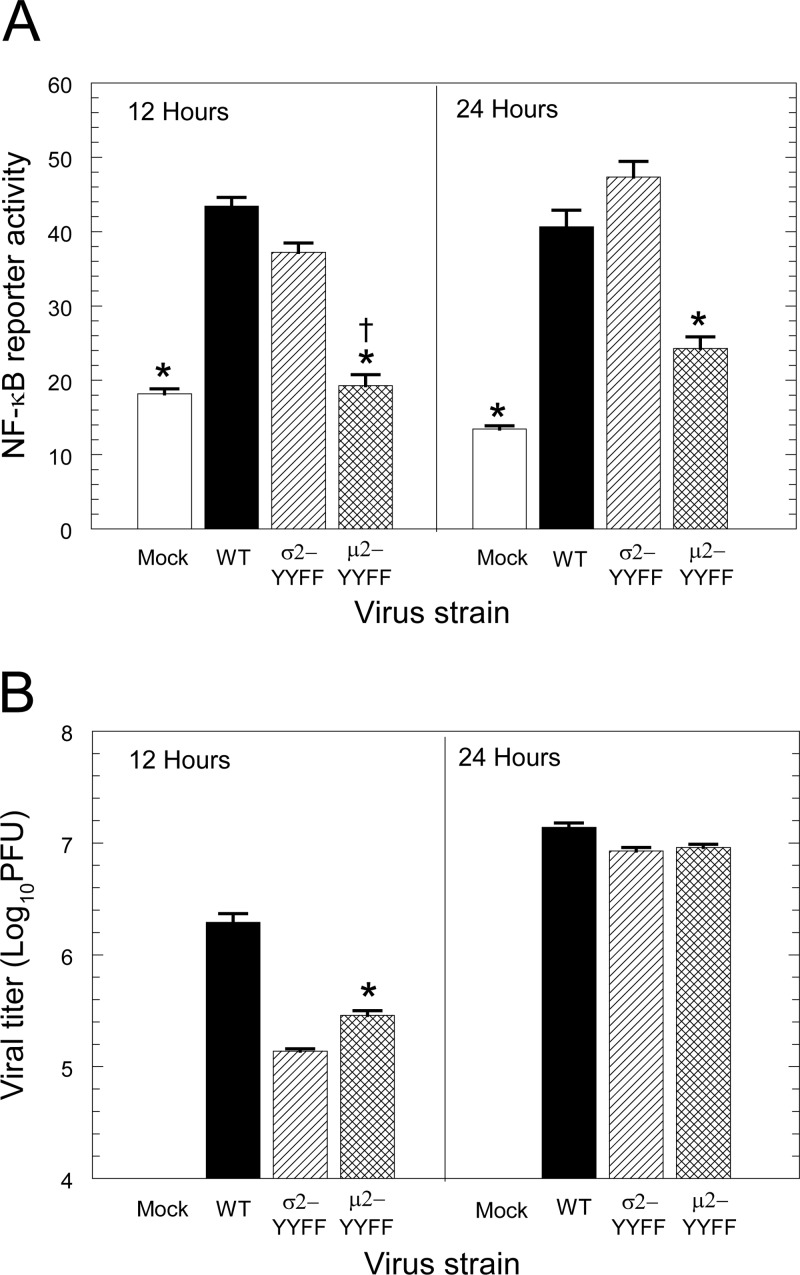
Immunoreceptor tyrosine-based activation motifs (ITAMs) are signaling domains located within the cytoplasmic tails of many transmembrane receptors and associated adaptor proteins that mediate immune cell activation. ITAMs also have been identified in the cytoplasmic tails of some enveloped virus glycoproteins. Here, we identified ITAM sequences in three mammalian reovirus proteins: μ2, σ2, and λ2. We demonstrate for the first time that μ2 is phosphorylated, contains a functional ITAM, and activates NF-κB. Specifically, μ2 and μNS recruit the ITAM-signaling intermediate Syk to cytoplasmic viral factories and this recruitment requires the μ2 ITAM. Moreover, both the μ2 ITAM and Syk are required for maximal μ2 activation of NF-κB. A mutant virus lacking the μ2 ITAM activates NF-κB less efficiently and induces lower levels of the downstream antiviral cytokine beta interferon (IFN-β) than does wild-type virus despite similar replication. Notably, the consequences of these μ2 ITAM effects are cell type specific. In fibroblasts where NF-κB is required for reovirus-induced apoptosis, the μ2 ITAM is advantageous for viral spread and enhances viral fitness. Conversely, in cardiac myocytes where the IFN response is critical for antiviral protection and NF-κB is not required for apoptosis, the μ2 ITAM stimulates cellular defense mechanisms and diminishes viral fitness. Together, these results suggest that the cell type-specific effect of the μ2 ITAM on viral spread reflects the cell type-specific effects of NF-κB and IFN-β. This first demonstration of a functional ITAM in a nonenveloped virus presents a new mechanism for viral ITAM-mediated signaling with likely organ-specific consequences in the host.
Figures







Similar articles
-
A single-amino-acid polymorphism in reovirus protein μ2 determines repression of interferon signaling and modulates myocarditis.J Virol. 2012 Feb;86(4):2302-11. doi: 10.1128/JVI.06236-11. Epub 2011 Dec 7. J Virol. 2012. PMID: 22156521 Free PMC article.
-
Downstream signaling molecules bind to different phosphorylated immunoreceptor tyrosine-based activation motif (ITAM) peptides of the high affinity IgE receptor.J Biol Chem. 1996 Nov 1;271(44):27962-8. doi: 10.1074/jbc.271.44.27962. J Biol Chem. 1996. PMID: 8910399
-
Differential Delivery of Genomic Double-Stranded RNA Causes Reovirus Strain-Specific Differences in Interferon Regulatory Factor 3 Activation.J Virol. 2018 Apr 13;92(9):e01947-17. doi: 10.1128/JVI.01947-17. Print 2018 May 1. J Virol. 2018. PMID: 29437975 Free PMC article.
-
C-type lectin receptor-induced NF-κB activation in innate immune and inflammatory responses.Cell Mol Immunol. 2012 Mar;9(2):105-12. doi: 10.1038/cmi.2011.58. Epub 2012 Jan 16. Cell Mol Immunol. 2012. PMID: 22246129 Free PMC article. Review.
-
Rotavirus and reovirus modulation of the interferon response.J Interferon Cytokine Res. 2009 Sep;29(9):559-67. doi: 10.1089/jir.2009.0072. J Interferon Cytokine Res. 2009. PMID: 19694545 Free PMC article. Review.
Cited by
-
Dissection of mammalian orthoreovirus µ2 reveals a self-associative domain required for binding to microtubules but not to factory matrix protein µNS.PLoS One. 2017 Sep 7;12(9):e0184356. doi: 10.1371/journal.pone.0184356. eCollection 2017. PLoS One. 2017. PMID: 28880890 Free PMC article.
-
A single amino acid substitution in the mRNA capping enzyme λ2 of a mammalian orthoreovirus mutant increases interferon sensitivity.Virology. 2015 Sep;483:229-35. doi: 10.1016/j.virol.2015.04.020. Epub 2015 May 15. Virology. 2015. PMID: 25985441 Free PMC article.
-
Cell Killing by Reovirus: Mechanisms and Consequences.Curr Top Microbiol Immunol. 2023;442:133-153. doi: 10.1007/82_2020_225. Curr Top Microbiol Immunol. 2023. PMID: 32986138 Free PMC article. Review.
-
Virophages and Their Interactions with Giant Viruses and Host Cells.Proteomes. 2018 May 22;6(2):23. doi: 10.3390/proteomes6020023. Proteomes. 2018. PMID: 29786634 Free PMC article. Review.
-
Loss of IKK Subunits Limits NF-κB Signaling in Reovirus-Infected Cells.J Virol. 2020 May 4;94(10):e00382-20. doi: 10.1128/JVI.00382-20. Print 2020 May 4. J Virol. 2020. PMID: 32161168 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 CA009385/CA/NCI NIH HHS/United States
- F32 AI075776/AI/NIAID NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- R01 AI50080/AI/NIAID NIH HHS/United States
- T32 CA09385/CA/NCI NIH HHS/United States
- R01 AI038296/AI/NIAID NIH HHS/United States
- CA68485/CA/NCI NIH HHS/United States
- R01 AI050080/AI/NIAID NIH HHS/United States
- R01 AI083333/AI/NIAID NIH HHS/United States
- R37 AI038296/AI/NIAID NIH HHS/United States
- DK20593/DK/NIDDK NIH HHS/United States
- R37 AI38296/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
