

Sequences related to SUMO interaction motifs in herpes simplex virus 1 protein ICP0 act cooperatively to stimulate virus infection
- PMID: 24352468
- PMCID: PMC3958091
- DOI: 10.1128/JVI.03417-13
Sequences related to SUMO interaction motifs in herpes simplex virus 1 protein ICP0 act cooperatively to stimulate virus infection
Abstract
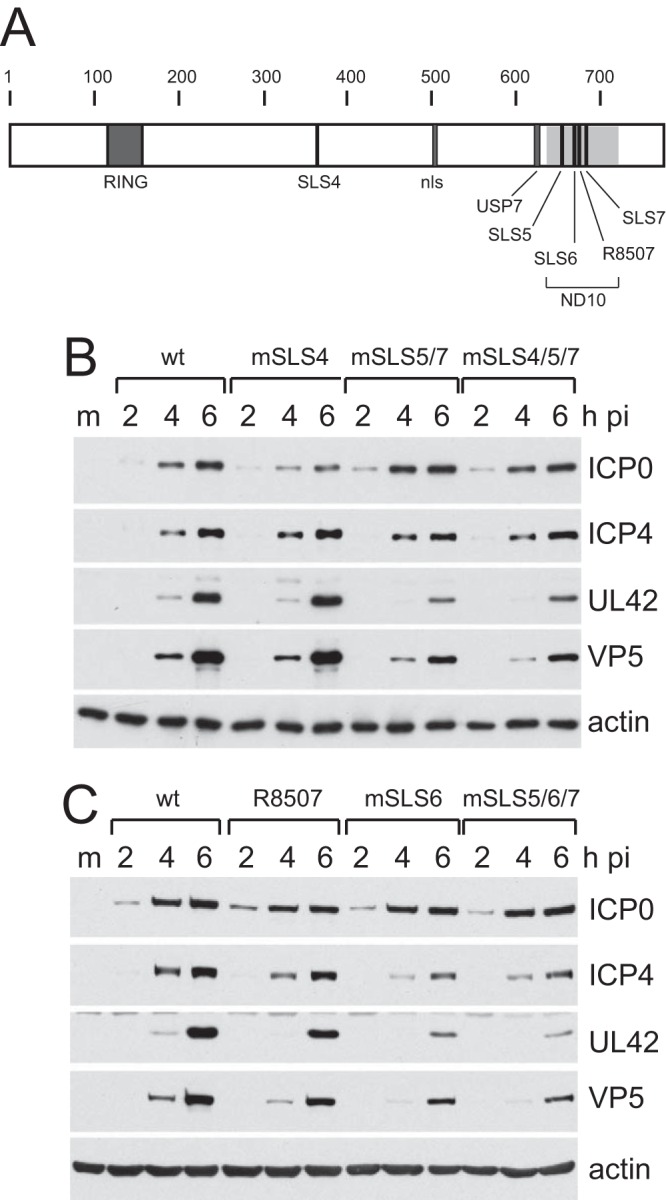
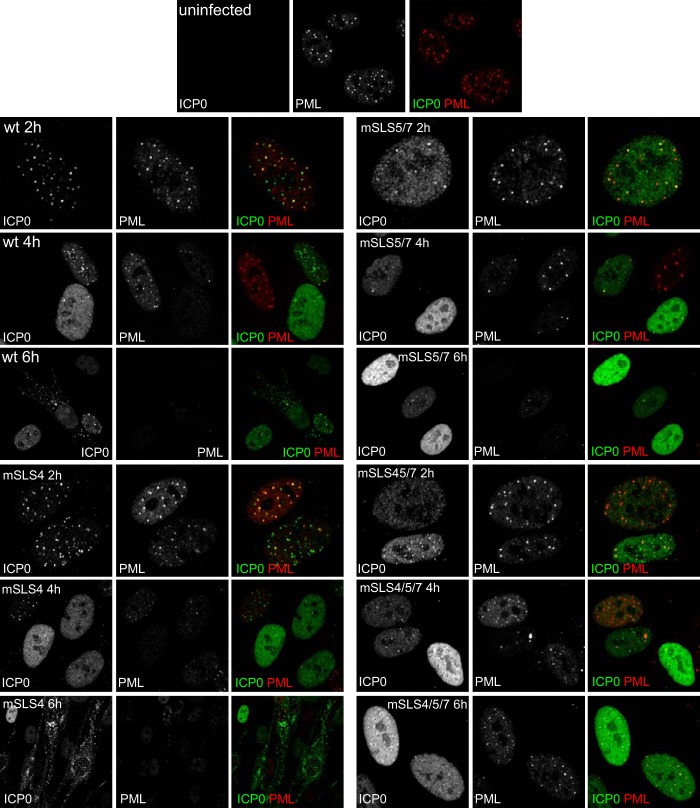
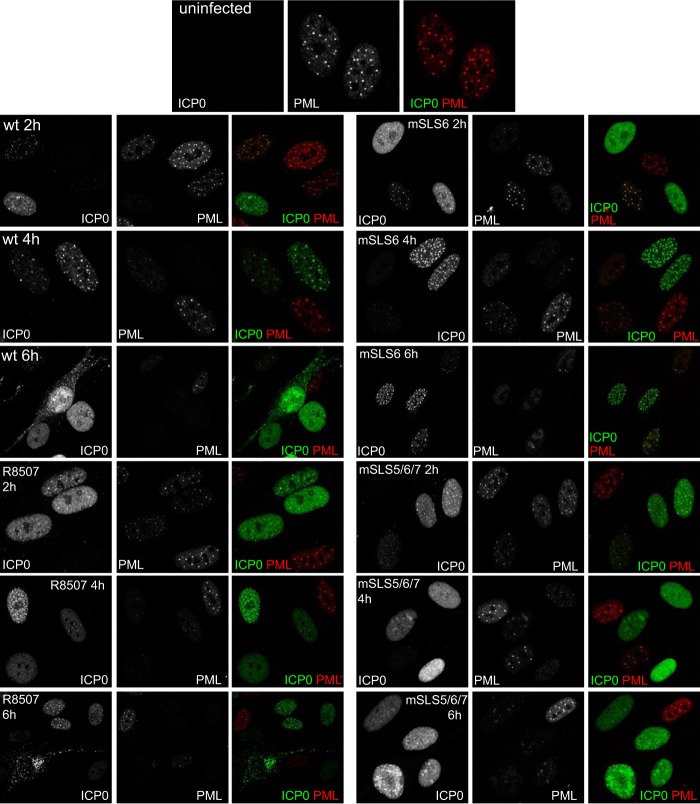
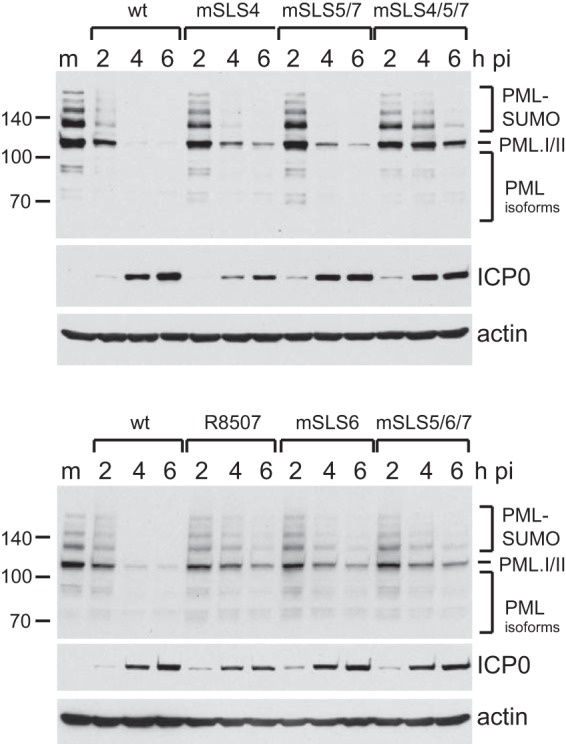
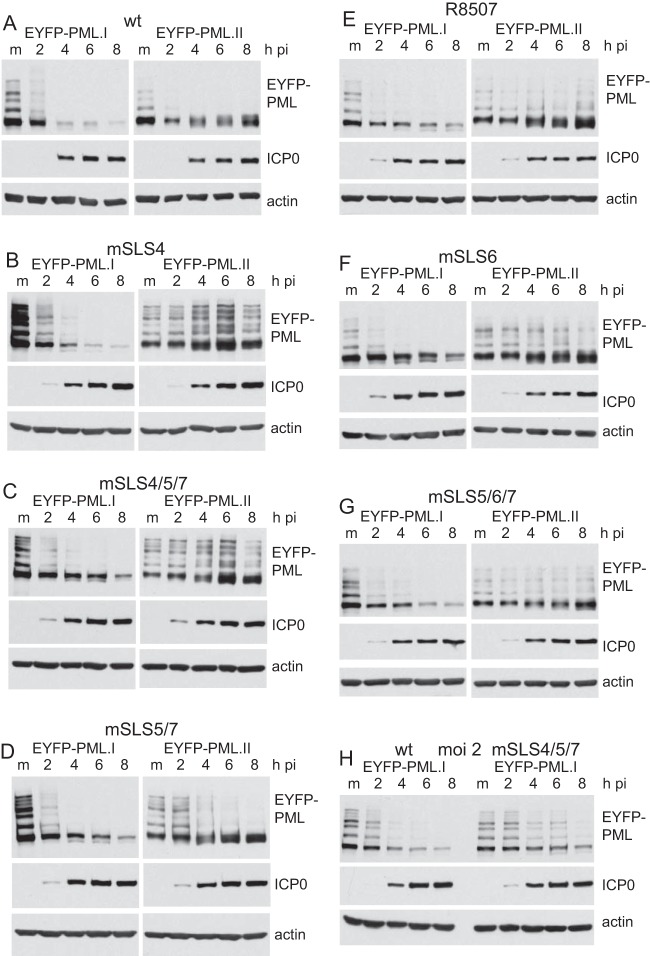
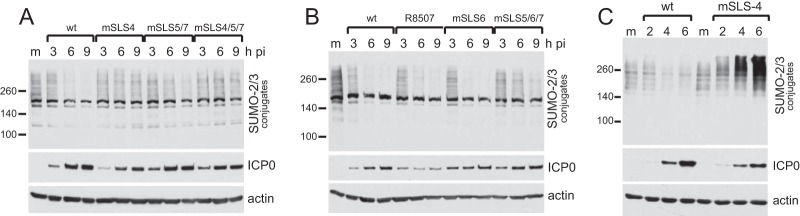
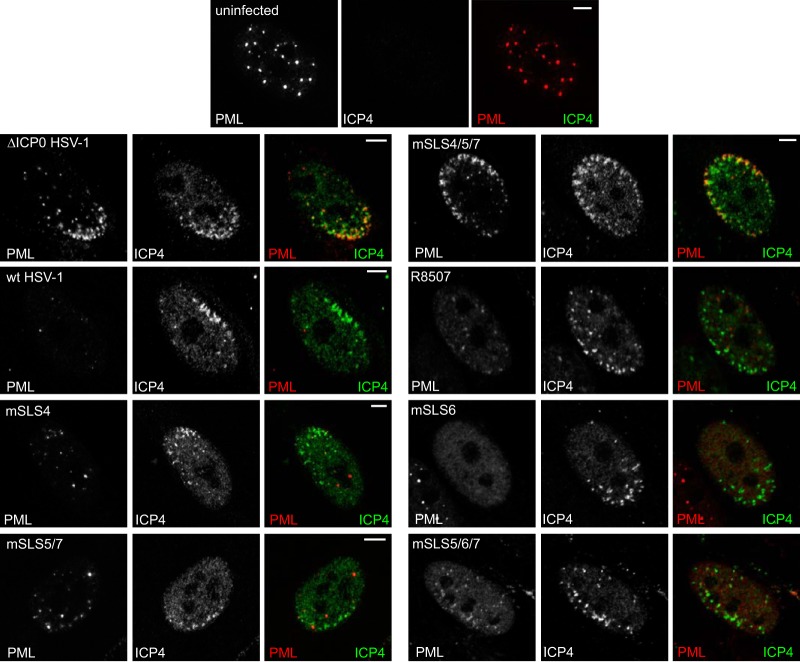
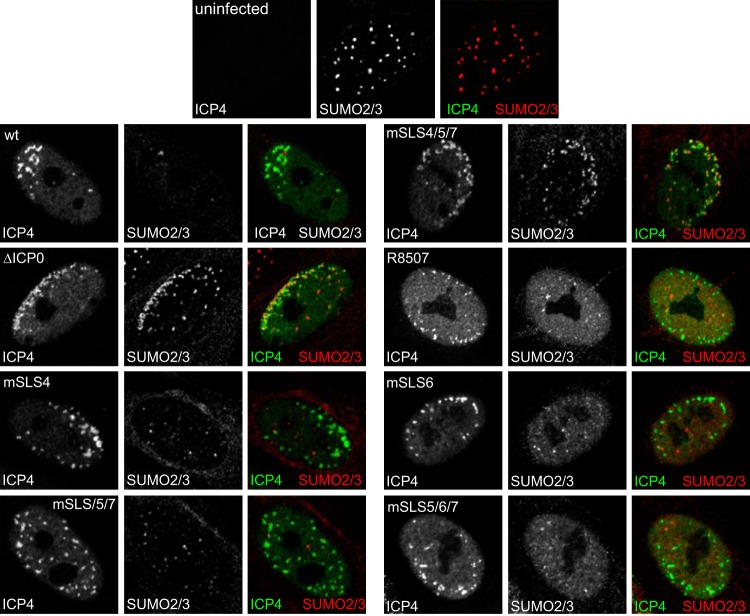
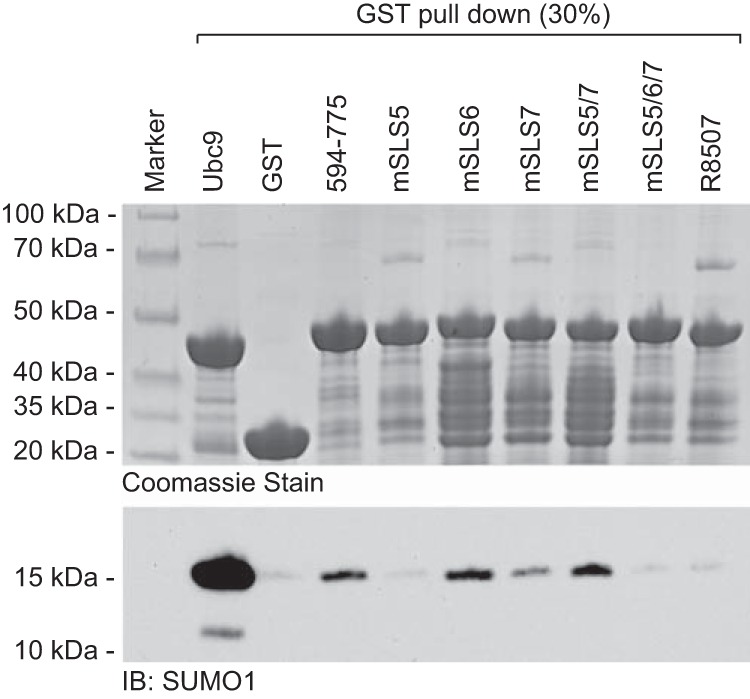
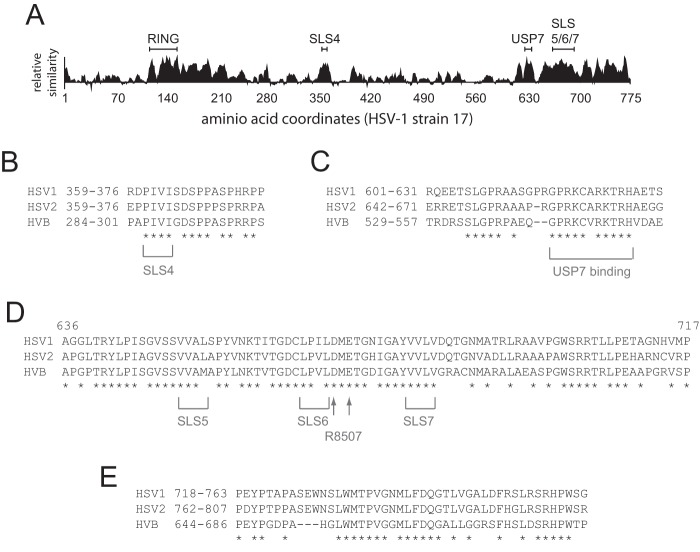
Herpes simplex virus type 1 immediate-early protein ICP0 is an E3 ubiquitin ligase of the RING finger class that degrades several cellular proteins during infection. This activity is essential for its functions in stimulating efficient lytic infection and productive reactivation from latency. ICP0 targets a number of proteins that are modified by the small ubiquitin-like SUMO family of proteins, and it includes a number of short sequences that are related to SUMO interaction motifs (SIMs). Therefore, ICP0 has characteristics that are related to those of cellular SUMO-targeted ubiquitin ligase enzymes. Here, we analyze the impact of mutation of a number of SIM-like sequences (SLSs) within ICP0 on HSV-1 replication and gene expression and their requirement for ICP0-mediated degradation of both sumoylated and unmodified promyelocytic leukemia (PML) and other sumoylated cellular proteins. One SLS in the central portion of the ICP0 sequence (SLS4) was found to be absolutely required for targeting cellular sumoylated species in general and sumoylated forms of PML other than those of PML isoform I. Mutation of a group of SLSs in the C-terminal quarter of ICP0 also reduced ICP0-mediated degradation of sumoylated PML in a cooperative manner. Although mutation of individual SLSs caused only modest decreases in viral replication, combined mutation of SLS4 with SLS sequences in the C-terminal quarter of the protein reduced plaque formation efficiency by up to two orders of magnitude. These results provide further evidence that the biological activities of ICP0 are connected with host cell sumoylation events.
Importance: Herpes simplex virus type 1 protein ICP0 plays important roles in regulating the initial stages of lytic infection and productive reactivation from latency. ICP0 mediates its effects through inducing the degradation of cellular proteins that have repressive effects on viral gene expression. An increasing number of cellular proteins are known to be sensitive to ICP0-mediated degradation; therefore, it is important to understand how ICP0 selects its substrates for degradation. This study identifies sequence motifs within ICP0 that are involved in targeting cellular proteins that are modified by the SUMO family of ubiquitin-like proteins and describes how mutation of combinations of these motifs causes a 100-fold defect in viral infectivity.
Figures
Similar articles
-
The HSV-1 ubiquitin ligase ICP0: Modifying the cellular proteome to promote infection.Virus Res. 2020 Aug;285:198015. doi: 10.1016/j.virusres.2020.198015. Epub 2020 May 13. Virus Res. 2020. PMID: 32416261 Free PMC article. Review.
-
Novel Role for Protein Inhibitor of Activated STAT 4 (PIAS4) in the Restriction of Herpes Simplex Virus 1 by the Cellular Intrinsic Antiviral Immune Response.J Virol. 2016 Apr 14;90(9):4807-4826. doi: 10.1128/JVI.03055-15. Print 2016 May. J Virol. 2016. PMID: 26937035 Free PMC article.
-
A viral ubiquitin ligase has substrate preferential SUMO targeted ubiquitin ligase activity that counteracts intrinsic antiviral defence.PLoS Pathog. 2011 Sep;7(9):e1002245. doi: 10.1371/journal.ppat.1002245. Epub 2011 Sep 15. PLoS Pathog. 2011. PMID: 21949651 Free PMC article.
-
Effect of SUMO-SIM Interaction on the ICP0-Mediated Degradation of PML Isoform II and Its Associated Proteins in Herpes Simplex Virus 1 Infection.J Virol. 2020 Jun 1;94(12):e00470-20. doi: 10.1128/JVI.00470-20. Print 2020 Jun 1. J Virol. 2020. PMID: 32295906 Free PMC article.
-
Regulation of alphaherpesvirus infections by the ICP0 family of proteins.J Gen Virol. 2013 Mar;94(Pt 3):465-481. doi: 10.1099/vir.0.048900-0. Epub 2012 Dec 12. J Gen Virol. 2013. PMID: 23239572 Review.
Cited by
-
Infected cell protein 0 functional domains and their coordination in herpes simplex virus replication.World J Virol. 2016 Feb 12;5(1):1-13. doi: 10.5501/wjv.v5.i1.1. World J Virol. 2016. PMID: 26870669 Free PMC article. Review.
-
The Role of ND10 Nuclear Bodies in Herpesvirus Infection: A Frenemy for the Virus?Viruses. 2021 Feb 3;13(2):239. doi: 10.3390/v13020239. Viruses. 2021. PMID: 33546431 Free PMC article. Review.
-
The HSV-1 ubiquitin ligase ICP0: Modifying the cellular proteome to promote infection.Virus Res. 2020 Aug;285:198015. doi: 10.1016/j.virusres.2020.198015. Epub 2020 May 13. Virus Res. 2020. PMID: 32416261 Free PMC article. Review.
-
A 77 Amino Acid Region in the N-Terminal Half of the HSV-1 E3 Ubiquitin Ligase ICP0 Contributes to Counteracting an Established Type 1 Interferon Response.Microbiol Spectr. 2022 Aug 31;10(4):e0059322. doi: 10.1128/spectrum.00593-22. Epub 2022 Jun 22. Microbiol Spectr. 2022. PMID: 35730940 Free PMC article.
-
A quantitative assay to monitor HSV-1 ICP0 ubiquitin ligase activity in vitro.Methods. 2015 Nov 15;90:3-7. doi: 10.1016/j.ymeth.2015.04.004. Epub 2015 Apr 8. Methods. 2015. PMID: 25862948 Free PMC article.
References
-
- Everett RD. 2011. The role of ICP0 in counteracting intrinsic cellular resistance to virus infection, p 39–50 In Weller SK. (ed), Alphaherpesviruses: molecular virology. Caister Academic Press, Norfolk, United Kingdom
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
