

The Role of Sdf-1α signaling in Xenopus laevis somite morphogenesis
- PMID: 24357195
- PMCID: PMC4040348
- DOI: 10.1002/dvdy.24092
The Role of Sdf-1α signaling in Xenopus laevis somite morphogenesis
Abstract
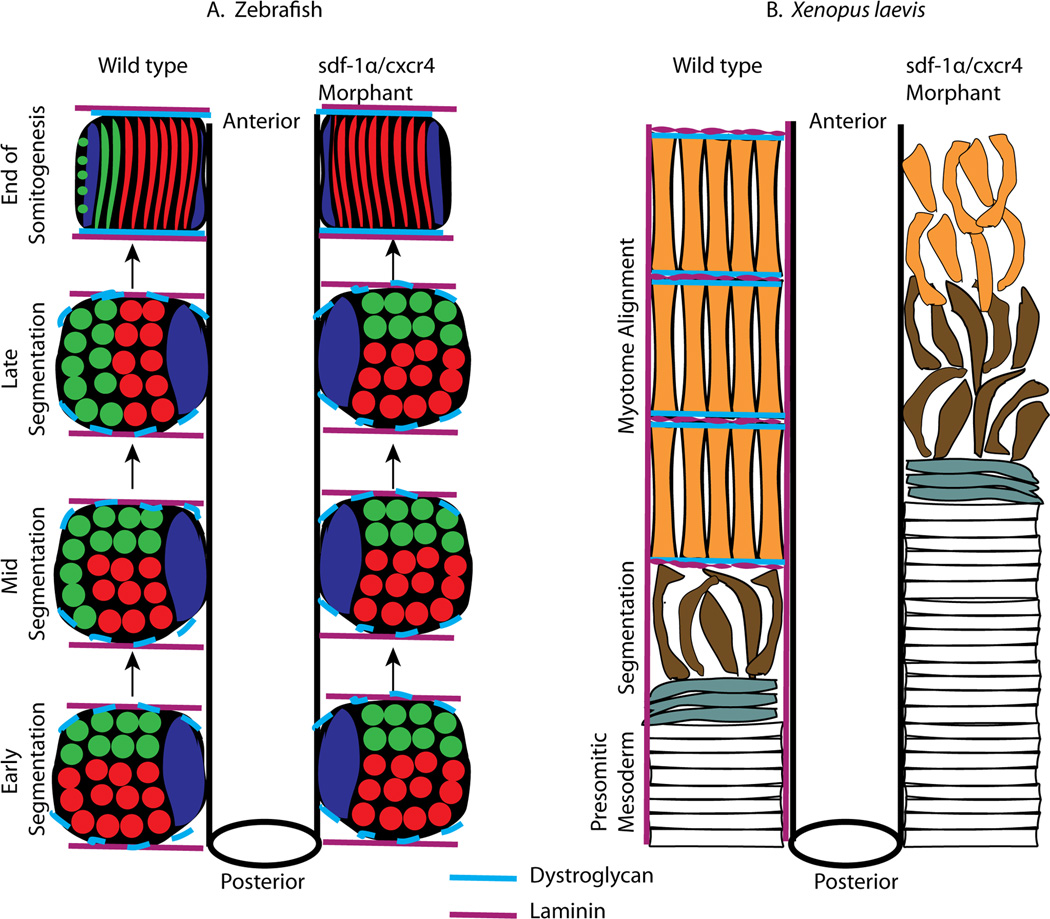
Background: Stromal derived factor-1α (sdf-1α), a chemoattractant chemokine, plays a major role in tumor growth, angiogenesis, metastasis, and in embryogenesis. The sdf-1α signaling pathway has also been shown to be important for somite rotation in zebrafish (Hollway et al., 2007). Given the known similarities and differences between zebrafish and Xenopus laevis somitogenesis, we sought to determine whether the role of sdf-1α is conserved in Xenopus laevis.
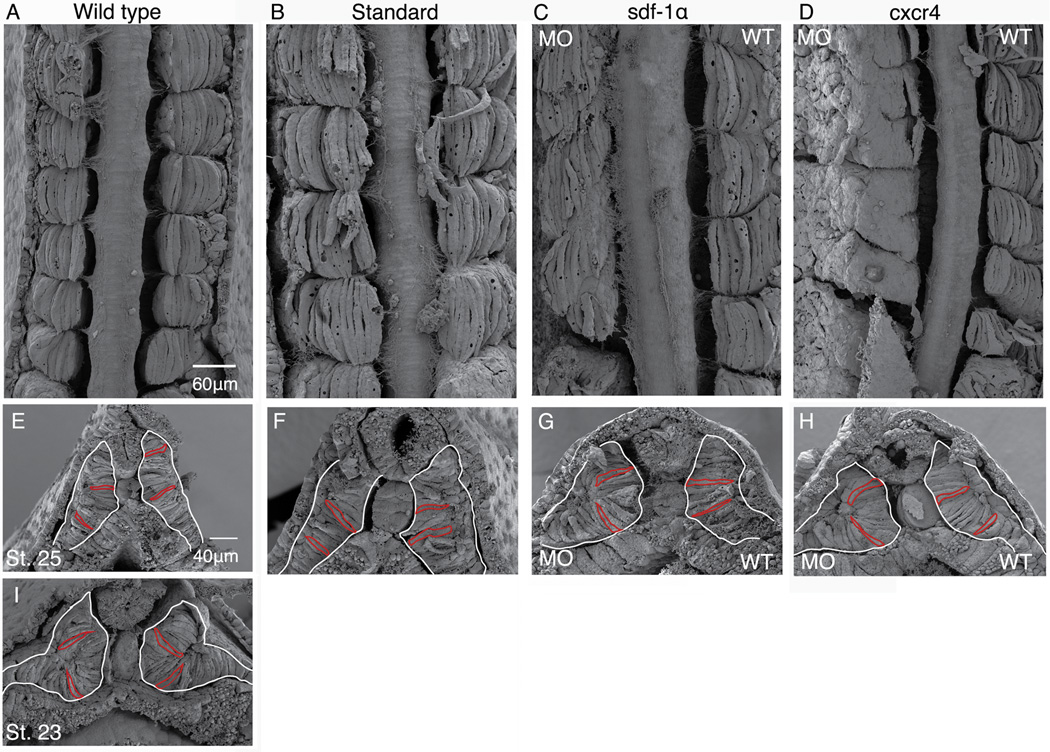
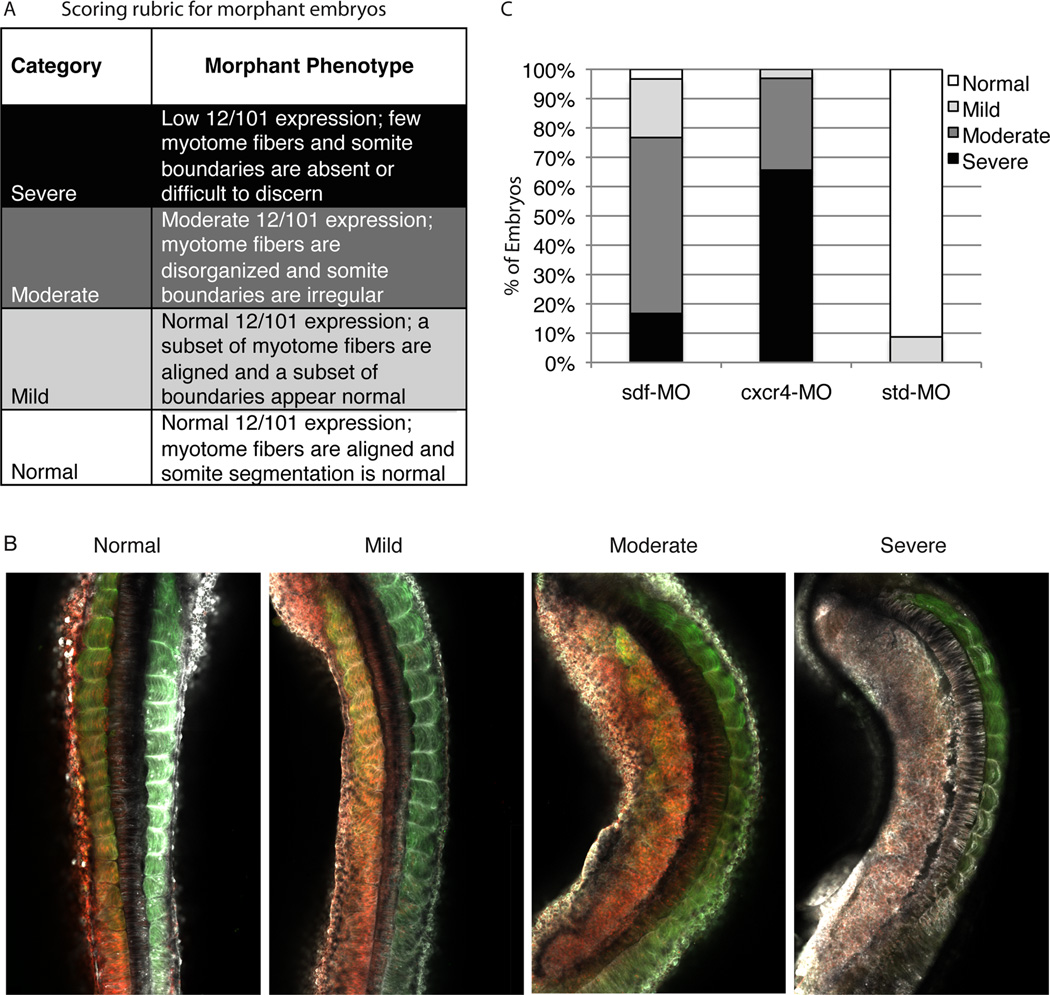
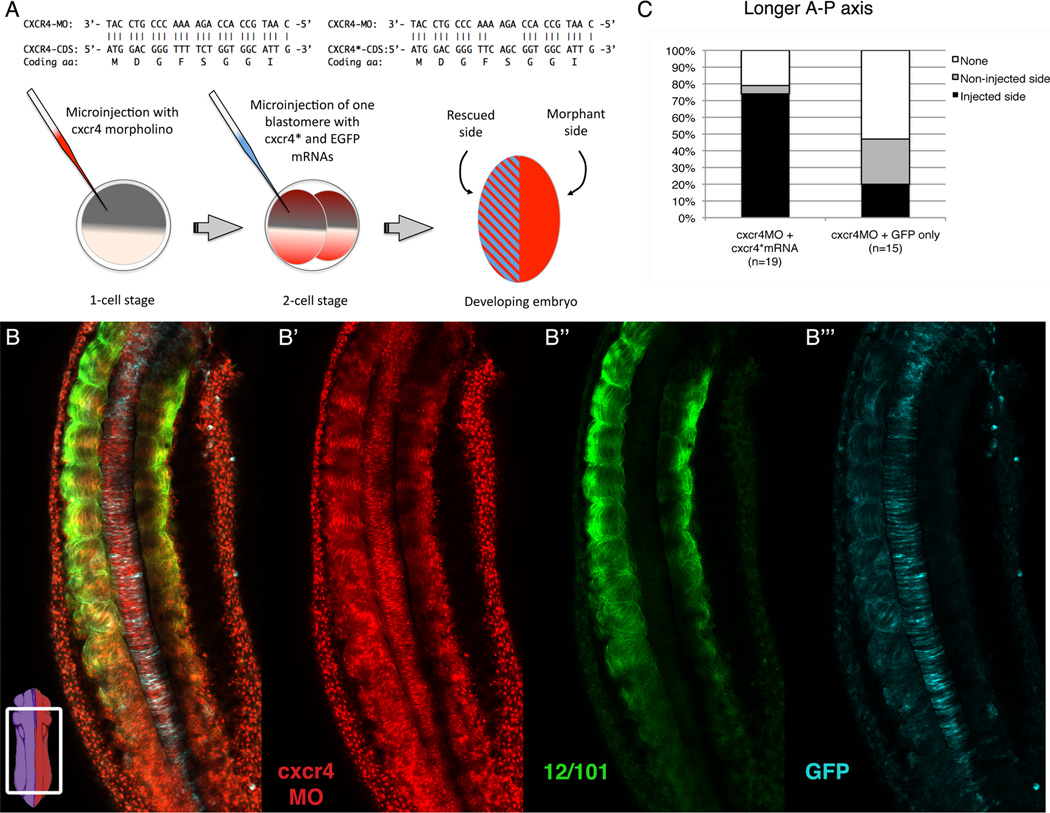
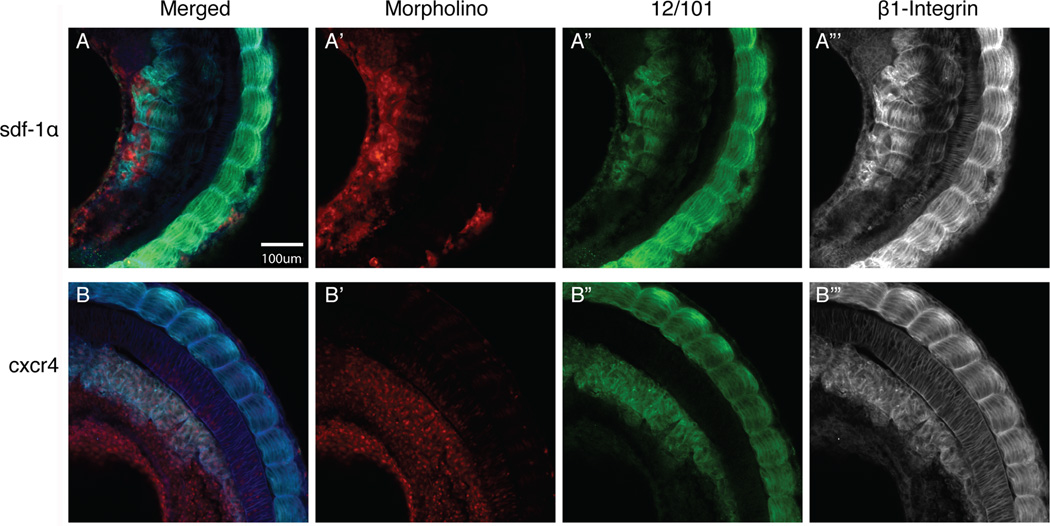
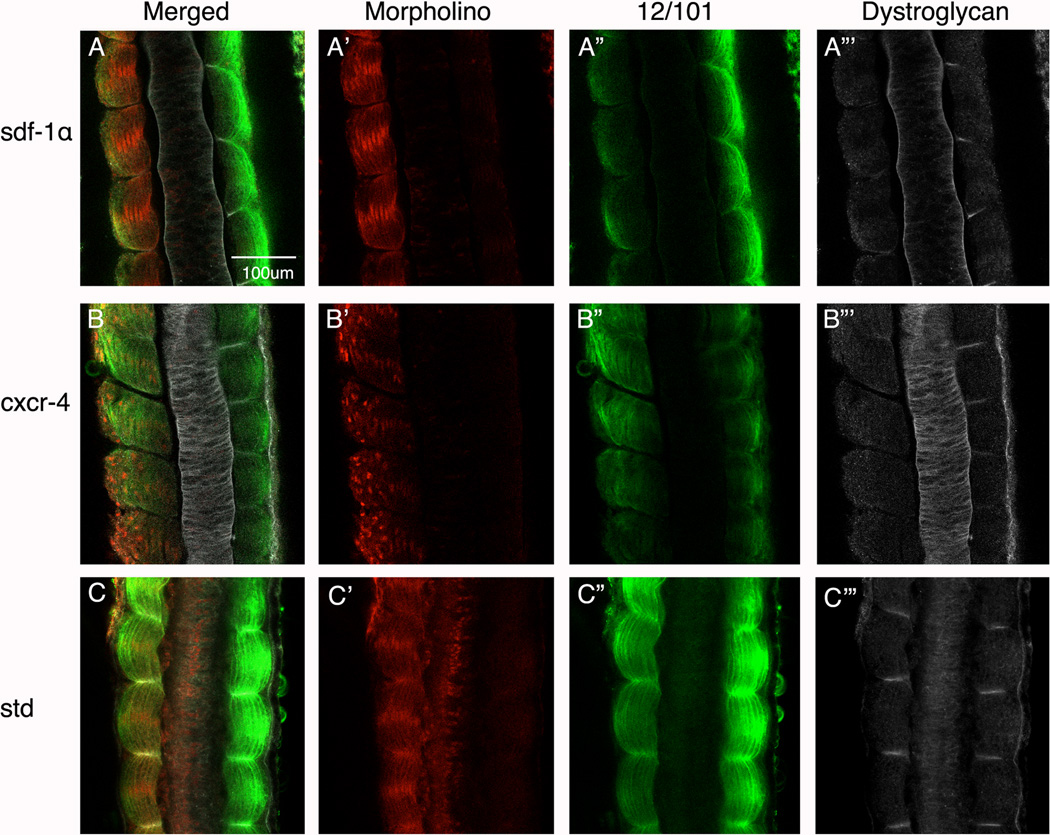
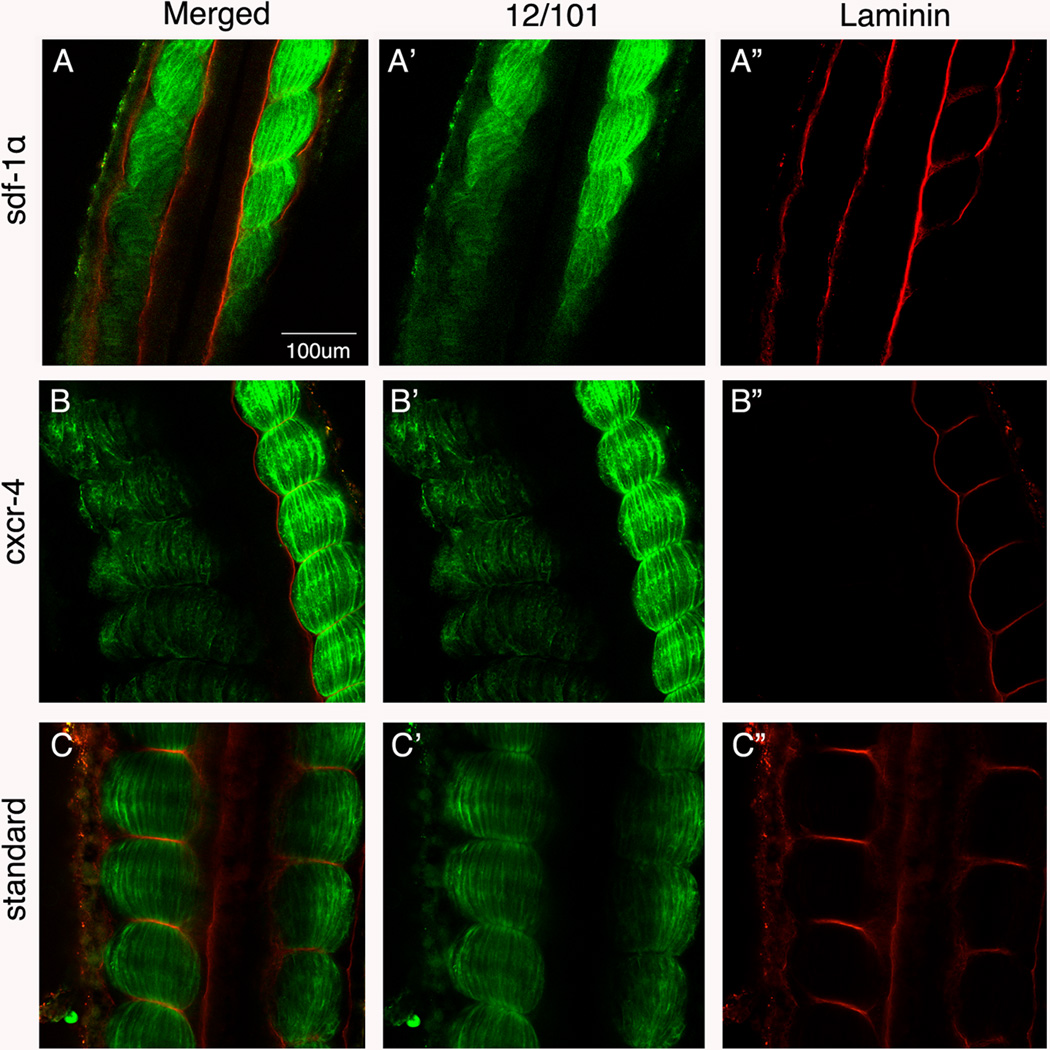
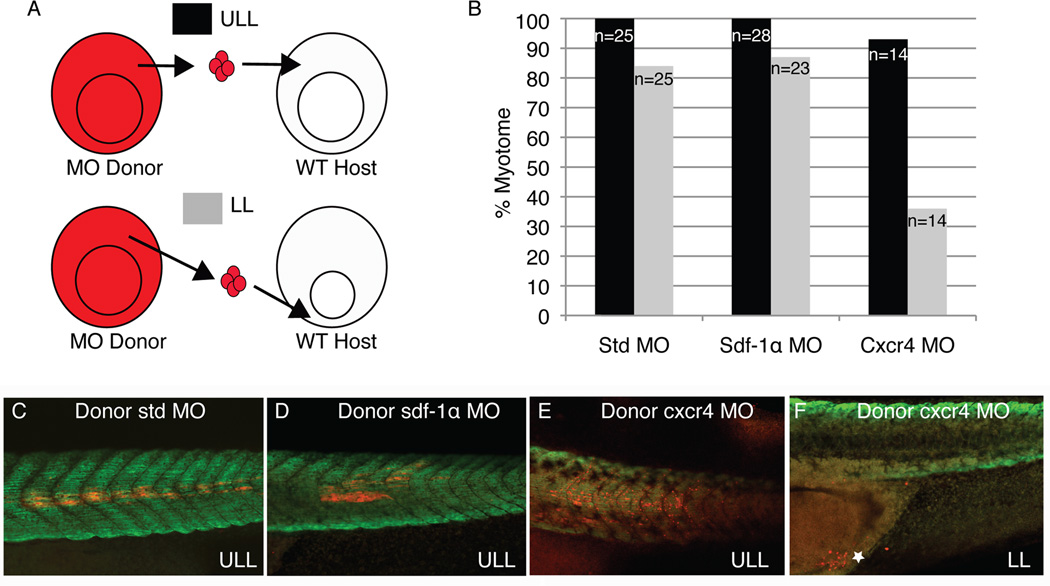
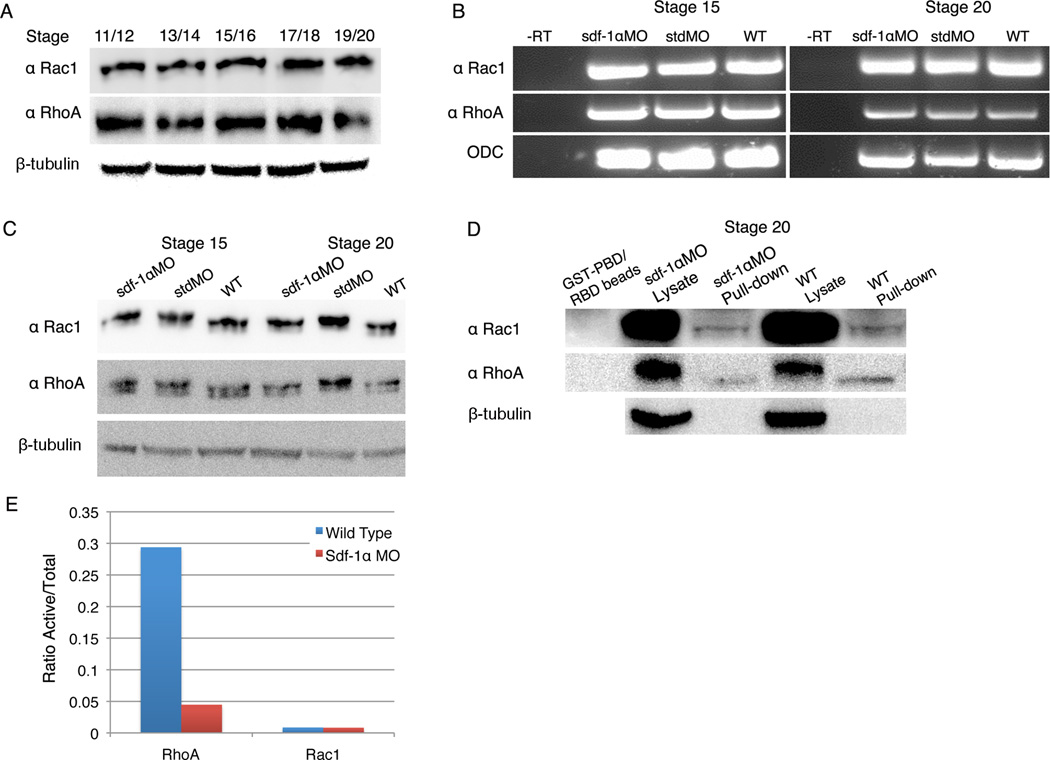
Results: Using a morpholino approach, we demonstrate that knockdown of sdf-1α or its receptor, cxcr4, leads to a significant disruption in somite rotation and myotome alignment. We further show that depletion of sdf-1α or cxcr4 leads to the near absence of β-dystroglycan and laminin expression at the intersomitic boundaries. Finally, knockdown of sdf-1α decreases the level of activated RhoA, a small GTPase known to regulate cell shape and movement.
Conclusion: Our results show that sdf-1α signaling regulates somite cell migration, rotation, and myotome alignment by directly or indirectly regulating dystroglycan expression and RhoA activation. These findings support the conservation of sdf-1α signaling in vertebrate somite morphogenesis; however, the precise mechanism by which this signaling pathway influences somite morphogenesis is different between the fish and the frog.
Keywords: RhoA; Xenopus laevis; cxcr4; morphogenesis; muscle; sdf-1α; somite; β-dystroglycan.
Copyright © 2013 Wiley Periodicals, Inc.
Figures
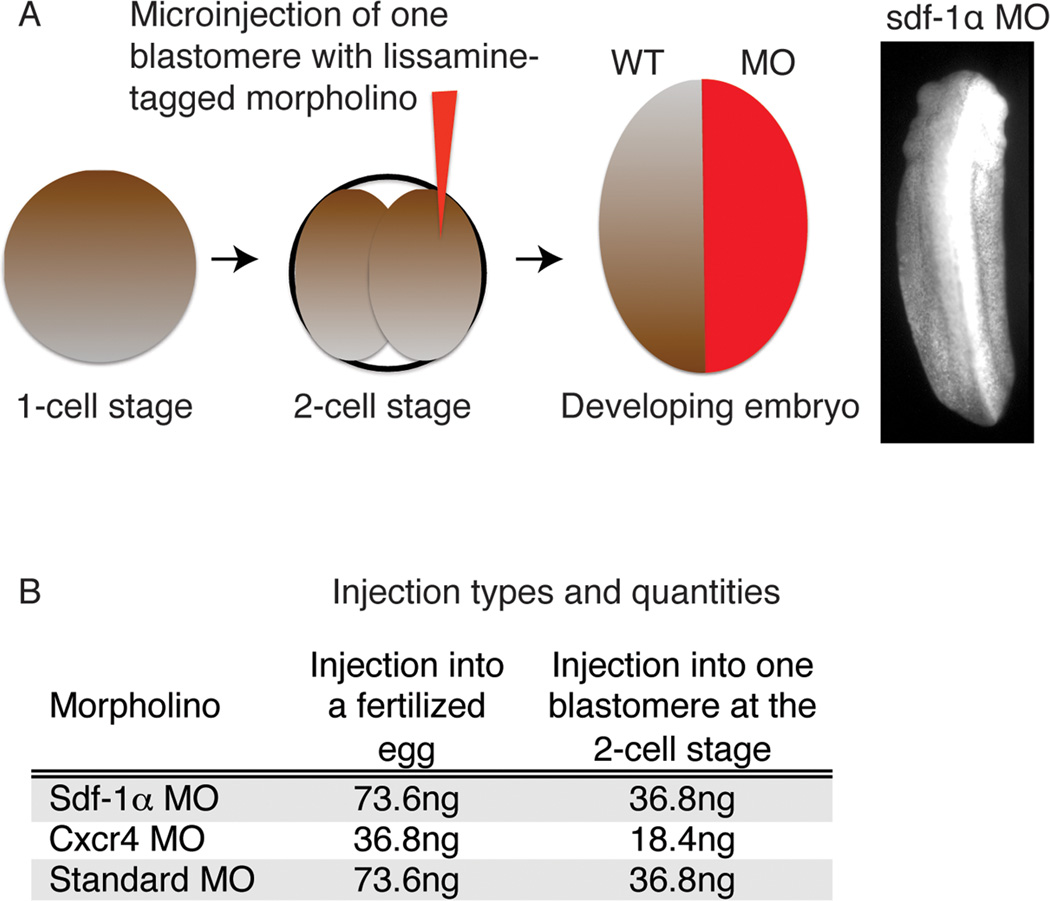
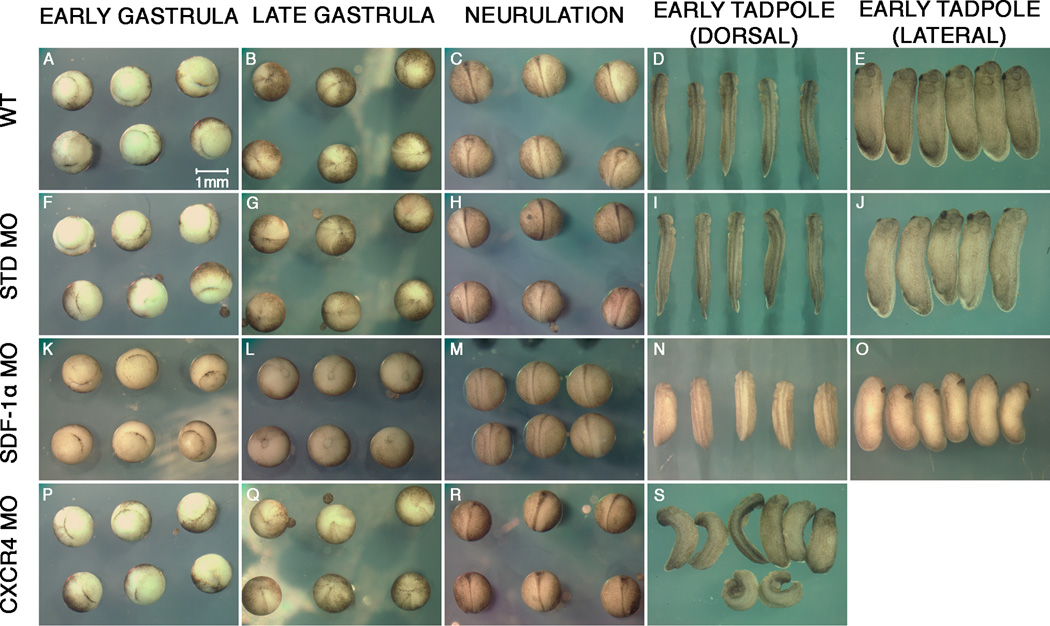
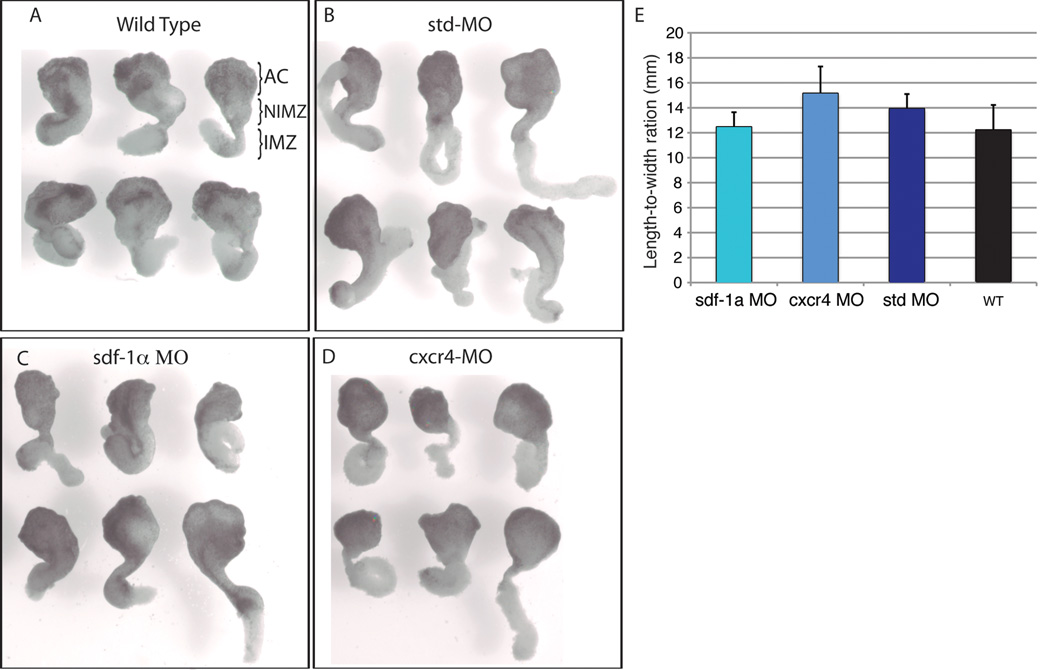
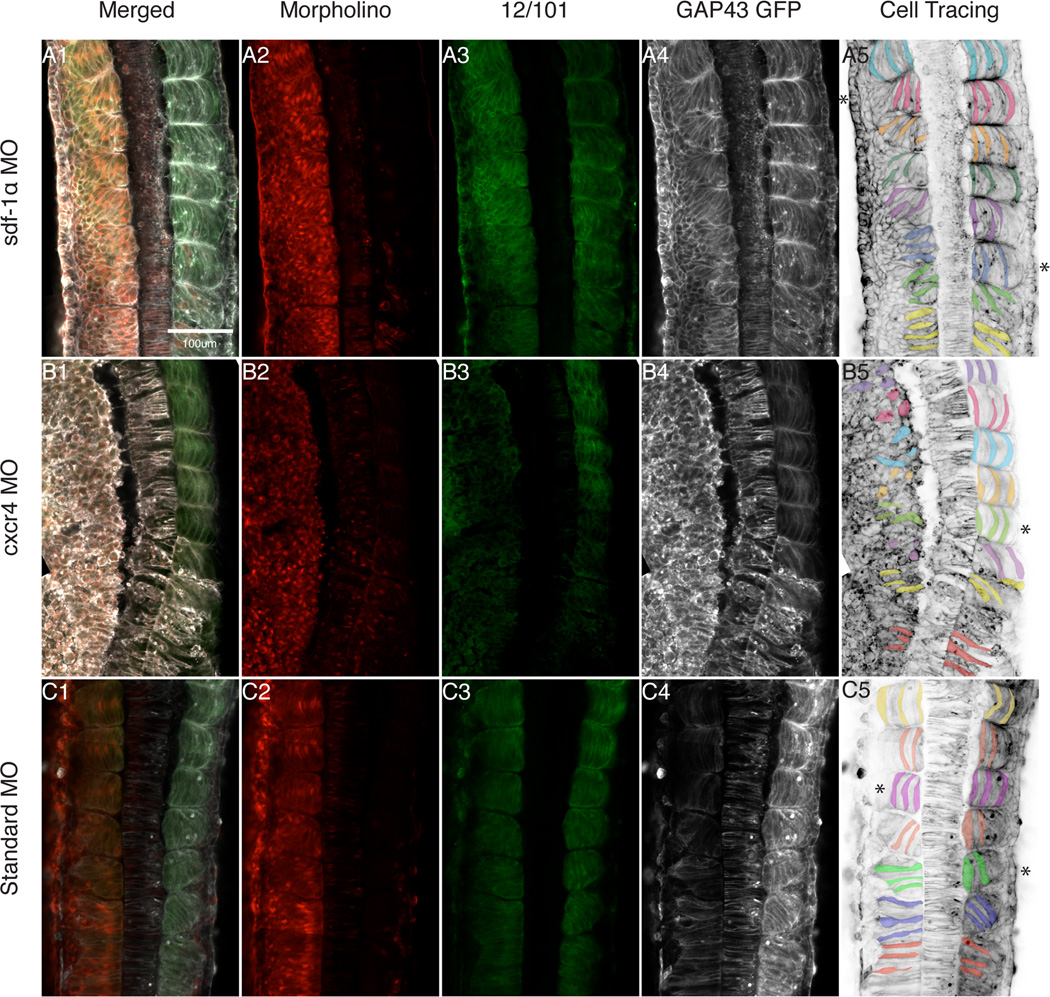
References
-
- Afonin B, Ho M, Gustin JK, Meloty-Kapella C, Domingo CR. Cell behaviors associated with somite segmentation and rotation in Xenopus laevis. Developmental Dynamics. 2006:3268–3279. - PubMed
-
- Bartolome R, Galvez B, Longo N, Baleux F, Van Muijen G, Sanchez-Mateos P, Arroyo A, Teixido J. Stromal Cell-Derived Factor-1α promotes melanoma cell invasion across basement membranes involving stimulation of membrane-type 1 matrix metalloproteinase and Rho GTPase activities. Cancer Res. 2004;64:2534–2543. - PubMed
-
- Benard V, Bohl BP, Bokoch GM. Characterization of Rac and Cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J Biol Chem. 1999;274:13198–13204. - PubMed
-
- Braun M, Wunderlin M, Spieth K, Knöchel W, Gierschik P, Moepps B. Xenopus laevis Stromal cell-derived factor 1: conservation of structure and function during vertebrate development. J Immuno. 2002;168(5):2340–2347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
