

Muscle acetylcholine receptor biosynthesis. Regulation by transcript availability
- PMID: 2435720
- PMCID: PMC5586536
Muscle acetylcholine receptor biosynthesis. Regulation by transcript availability
Abstract
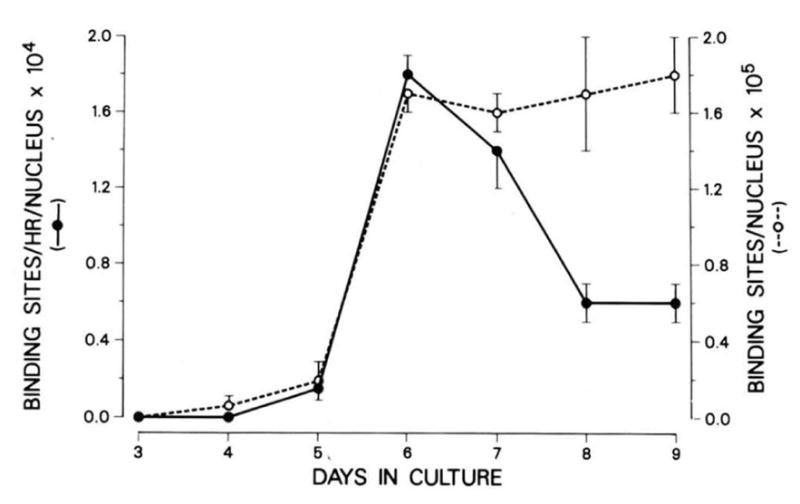
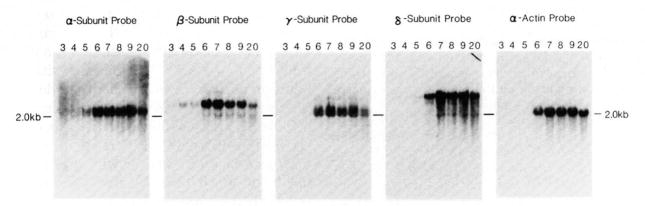
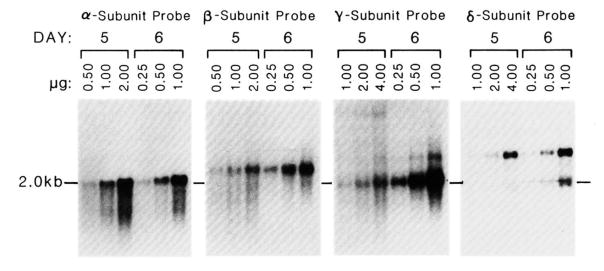
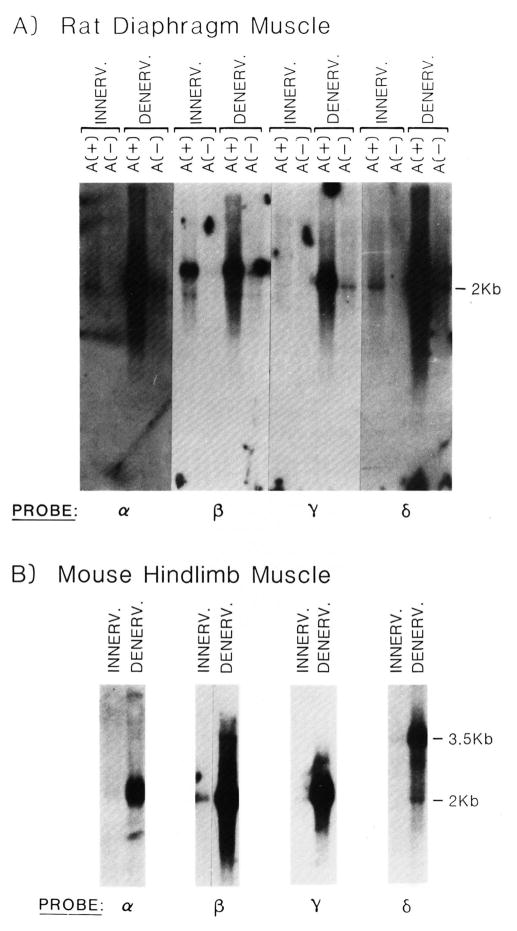
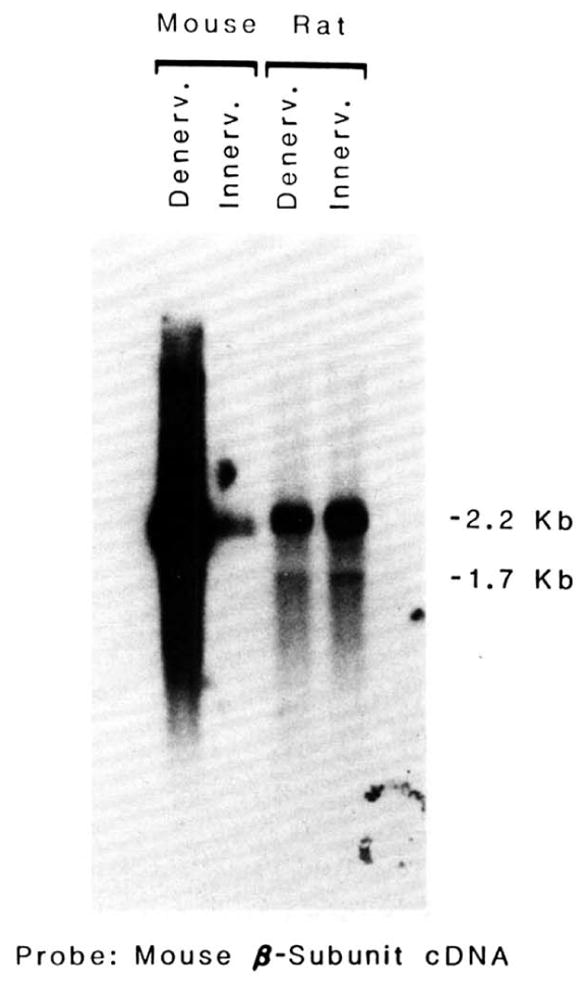
The expression of the muscle nicotinic acetylcholine receptor (AChR) on the cell surface entails a complex biosynthetic pathway, involving the expression and assembly of four subunits. The amount of AChR on the cell surface changes throughout muscle development and upon muscle denervation. We have examined the regulatory role of transcript levels on surface AChR expression by RNA blot analysis. During myogenesis of the fusing mouse muscle cell line C2, which expresses an embryonic type of receptor, changes in the rate of appearance of cell surface AChR have been assayed by 125I-alpha-bungarotoxin binding. The maximal increase in the rate of appearance of cell surface AChR occurs upon cell fusion, closely following the maximal increase in transcript levels for the alpha-, beta-, gamma-, and delta-AChR subunits. AChR alpha-, beta-, gamma-, and delta-subunit gene transcript levels have also been examined in innervated and denervated rat and mouse muscle. Muscle denervation results in an increase of transcripts for all four subunits. However, the amount of beta-subunit transcript in innervated rat skeletal muscle is high relative to the other subunit transcripts, and increases less than the other subunit transcripts upon denervation. Our results indicate that, during myogenesis and upon denervation, the availability of AChR subunit transcripts for translation plays a regulatory role in surface AChR appearance.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
