

The oxidative burst reaction in mammalian cells depends on gravity
- PMID: 24359439
- PMCID: PMC3880029
- DOI: 10.1186/1478-811X-11-98
The oxidative burst reaction in mammalian cells depends on gravity
Abstract
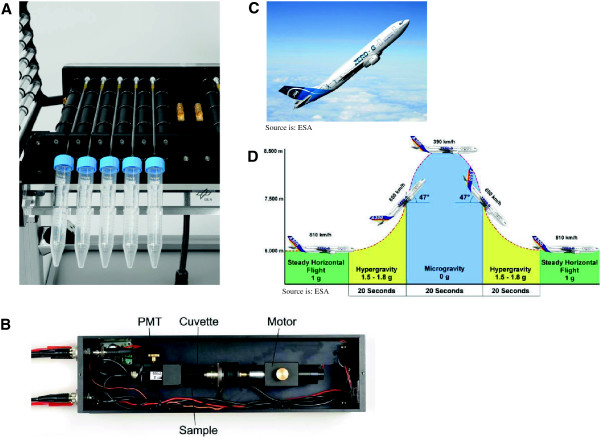
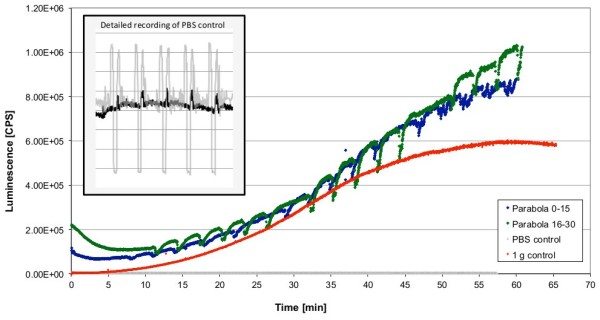
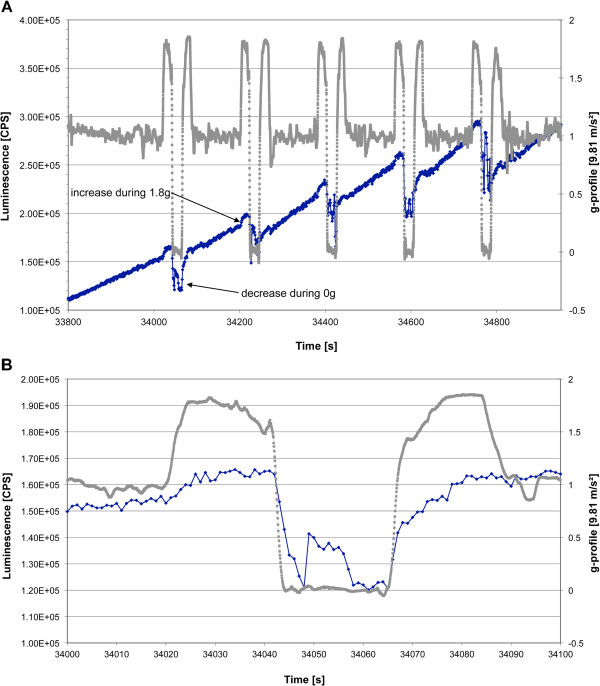
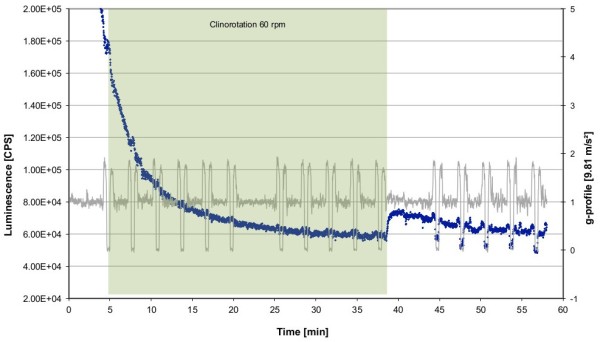
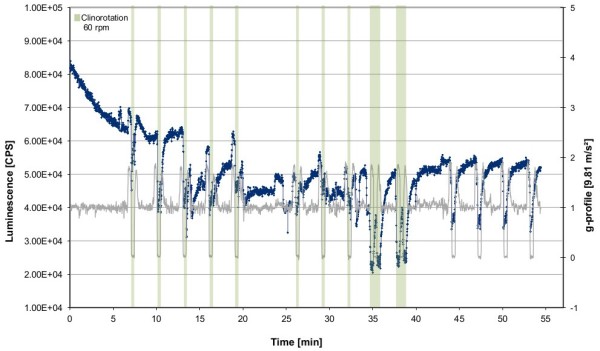
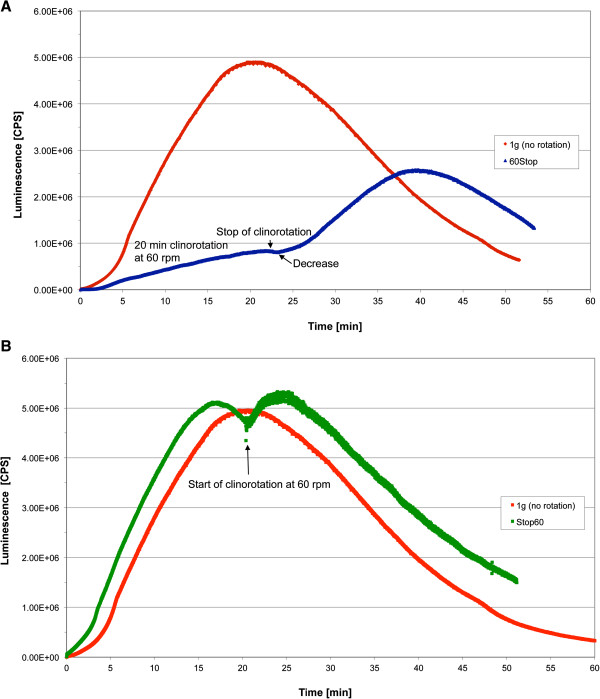
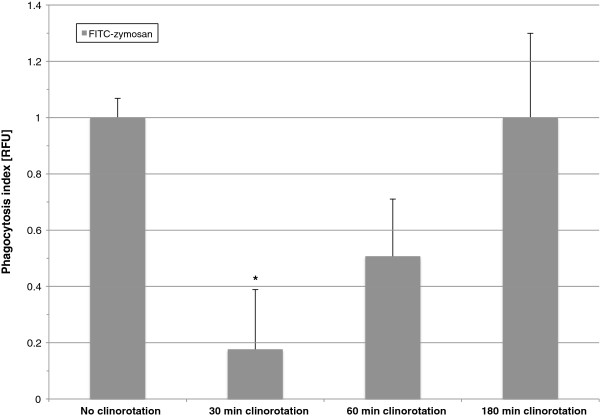
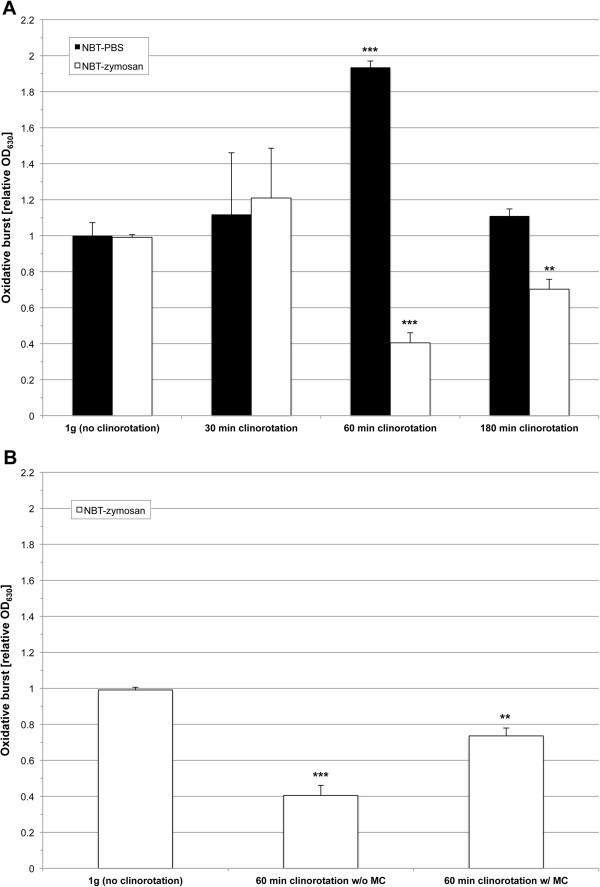
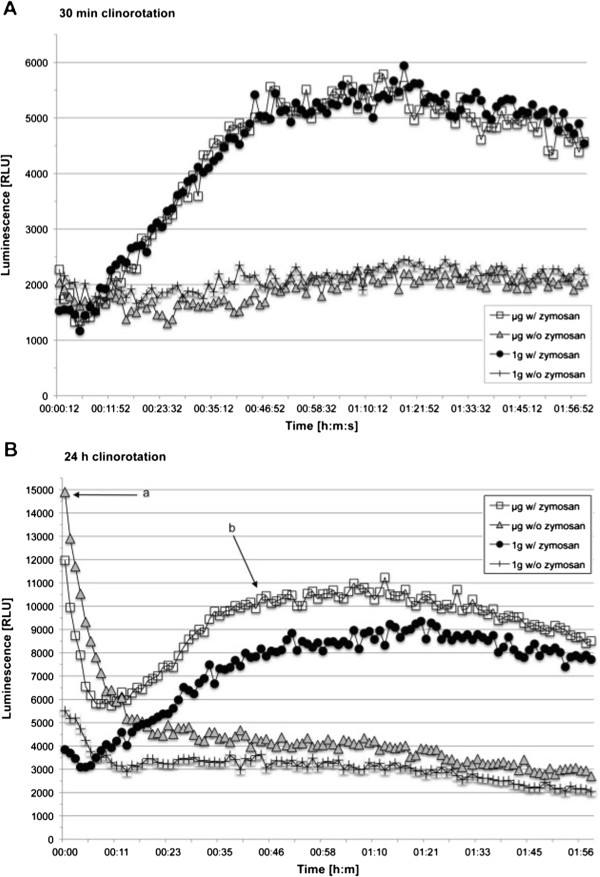
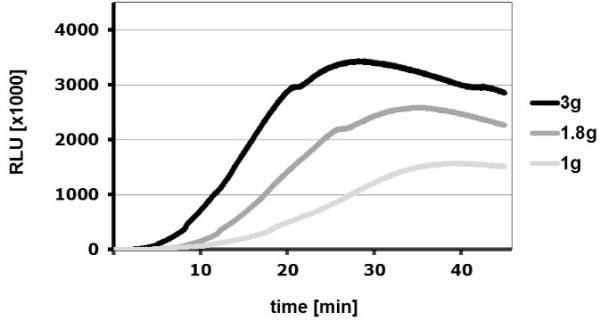
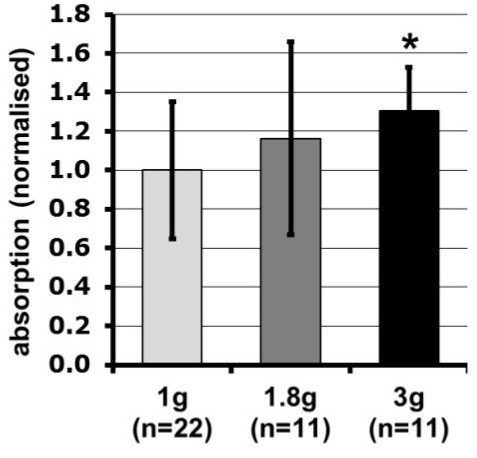
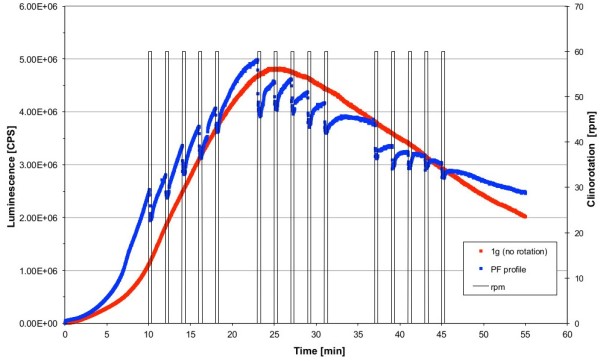
Gravity has been a constant force throughout the Earth's evolutionary history. Thus, one of the fundamental biological questions is if and how complex cellular and molecular functions of life on Earth require gravity. In this study, we investigated the influence of gravity on the oxidative burst reaction in macrophages, one of the key elements in innate immune response and cellular signaling. An important step is the production of superoxide by the NADPH oxidase, which is rapidly converted to H2O2 by spontaneous and enzymatic dismutation. The phagozytosis-mediated oxidative burst under altered gravity conditions was studied in NR8383 rat alveolar macrophages by means of a luminol assay. Ground-based experiments in "functional weightlessness" were performed using a 2 D clinostat combined with a photomultiplier (PMT clinostat). The same technical set-up was used during the 13th DLR and 51st ESA parabolic flight campaign. Furthermore, hypergravity conditions were provided by using the Multi-Sample Incubation Centrifuge (MuSIC) and the Short Arm Human Centrifuge (SAHC). The results demonstrate that release of reactive oxygen species (ROS) during the oxidative burst reaction depends greatly on gravity conditions. ROS release is 1.) reduced in microgravity, 2.) enhanced in hypergravity and 3.) responds rapidly and reversible to altered gravity within seconds. We substantiated the effect of altered gravity on oxidative burst reaction in two independent experimental systems, parabolic flights and 2D clinostat / centrifuge experiments. Furthermore, the results obtained in simulated microgravity (2D clinorotation experiments) were proven by experiments in real microgravity as in both cases a pronounced reduction in ROS was observed. Our experiments indicate that gravity-sensitive steps are located both in the initial activation pathways and in the final oxidative burst reaction itself, which could be explained by the role of cytoskeletal dynamics in the assembly and function of the NADPH oxidase complex.
Figures
References
-
- Cogoli A. Gravitational physiology of human immune cells: a review of in vivo, ex vivo and in vitro studies. J Gravit Physiol. 1996;11(1):1–9. - PubMed
-
- Bräucker R, Cogoli A, Hemmersbach R. In: Astrobiology: The Quest for the Conditions of Life. Baumstark-Khan C, Horneck GG, editor. Berlin: Heidelberg: Springer; 2001. Graviperception and graviresponse at the cellular level; pp. 284–297.
-
- Tauber S, Hauschild S, Crescio C, Secchi C, Paulsen K, Pantaleo A, Saba A, Buttron I, Thiel CS, Cogoli A, Pippia P, Ullrich O. Signal transduction in primary human T lymphocytes in altered gravity -results of the MASER-12 suborbital space flight mission. Cell Commun Signal. 2013;11(1):32. doi: 10.1186/1478-811X-11-32. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
