

Direct glucose production from lignocellulose using Clostridium thermocellum cultures supplemented with a thermostable β-glucosidase
- PMID: 24359557
- PMCID: PMC3878107
- DOI: 10.1186/1754-6834-6-184
Direct glucose production from lignocellulose using Clostridium thermocellum cultures supplemented with a thermostable β-glucosidase
Abstract
Background: Cellulases continue to be one of the major costs associated with the lignocellulose hydrolysis process. Clostridium thermocellum is an anaerobic, thermophilic, cellulolytic bacterium that produces cellulosomes capable of efficiently degrading plant cell walls. The end-product cellobiose, however, inhibits degradation. To maximize the cellulolytic ability of C. thermocellum, it is important to eliminate this end-product inhibition.
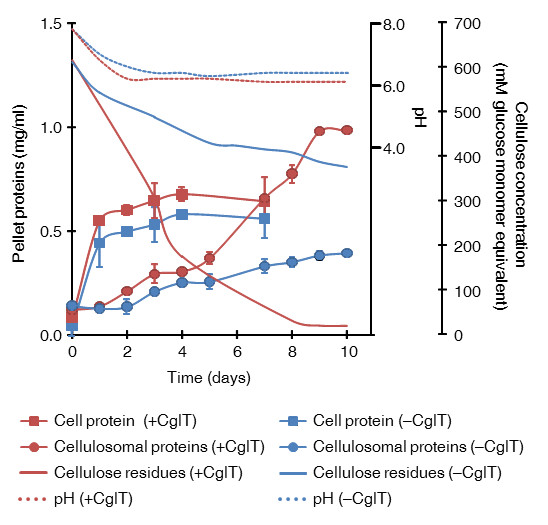
Results: This work describes a system for biological saccharification that leads to glucose production following hydrolysis of lignocellulosic biomass. C. thermocellum cultures supplemented with thermostable beta-glucosidases make up this system. This approach does not require any supplementation with cellulases and hemicellulases. When C. thermocellum strain S14 was cultured with a Thermoanaerobacter brockii beta-glucosidase (CglT with activity 30 U/g cellulose) in medium containing 100 g/L cellulose (617 mM initial glucose equivalents), we observed not only high degradation of cellulose, but also accumulation of 426 mM glucose in the culture broth. In contrast, cultures without CglT, or with less thermostable beta-glucosidases, did not efficiently hydrolyze cellulose and accumulated high levels of glucose. Glucose production required a cellulose load of over 10 g/L. When alkali-pretreated rice straw containing 100 g/L glucan was used as the lignocellulosic biomass, approximately 72% of the glucan was saccharified, and glucose accumulated to 446 mM in the culture broth. The hydrolysate slurry containing glucose was directly fermented to 694 mM ethanol by addition of Saccharomyces cerevisiae, giving an 85% theoretical yield without any inhibition.
Conclusions: Our process is the first instance of biological saccharification with exclusive production and accumulation of glucose from lignocellulosic biomass. The key to its success was the use of C. thermocellum supplemented with a thermostable beta-glucosidase and cultured under a high cellulose load. We named this approach biological simultaneous enzyme production and saccharification (BSES). BSES may resolve a significant barrier to economical production by providing a platform for production of fermentable sugars with reduced enzyme amounts.
Figures





Similar articles
-
Efficient saccharification of ammonia soaked rice straw by combination of Clostridium thermocellum cellulosome and Thermoanaerobacter brockii β-glucosidase.Bioresour Technol. 2012 Mar;107:352-7. doi: 10.1016/j.biortech.2011.12.126. Epub 2012 Jan 2. Bioresour Technol. 2012. PMID: 22257861
-
Lignocellulosic saccharification by a newly isolated bacterium, Ruminiclostridium thermocellum M3 and cellular cellulase activities for high ratio of glucose to cellobiose.Biotechnol Biofuels. 2016 Aug 11;9:172. doi: 10.1186/s13068-016-0585-z. eCollection 2016. Biotechnol Biofuels. 2016. PMID: 27525041 Free PMC article.
-
Glucose production from cellulose through biological simultaneous enzyme production and saccharification using recombinant bacteria expressing the β-glucosidase gene.J Biosci Bioeng. 2019 Mar;127(3):340-344. doi: 10.1016/j.jbiosc.2018.08.008. Epub 2018 Sep 17. J Biosci Bioeng. 2019. PMID: 30237013
-
Intracellular cellobiose metabolism and its applications in lignocellulose-based biorefineries.Bioresour Technol. 2017 Sep;239:496-506. doi: 10.1016/j.biortech.2017.05.001. Epub 2017 May 4. Bioresour Technol. 2017. PMID: 28535986 Review.
-
Bioconversion of lignocellulosic biomass: biochemical and molecular perspectives.J Ind Microbiol Biotechnol. 2008 May;35(5):377-391. doi: 10.1007/s10295-008-0327-8. Epub 2008 Mar 13. J Ind Microbiol Biotechnol. 2008. PMID: 18338189 Review.
Cited by
-
Metabolome Analysis of Constituents in Membrane Vesicles for Clostridium thermocellum Growth Stimulation.Microorganisms. 2021 Mar 13;9(3):593. doi: 10.3390/microorganisms9030593. Microorganisms. 2021. PMID: 33805707 Free PMC article.
-
Artificial consortium demonstrates emergent properties of enhanced cellulosic-sugar degradation and biofuel synthesis.NPJ Biofilms Microbiomes. 2020 Dec 2;6(1):59. doi: 10.1038/s41522-020-00170-8. NPJ Biofilms Microbiomes. 2020. PMID: 33268782 Free PMC article.
-
Material utilization of green waste: a review on potential valorization methods.Bioresour Bioprocess. 2021 Feb 22;8(1):19. doi: 10.1186/s40643-021-00367-5. Bioresour Bioprocess. 2021. PMID: 38650228 Free PMC article. Review.
-
Efficient whole-cell-catalyzing cellulose saccharification using engineered Clostridium thermocellum.Biotechnol Biofuels. 2017 May 12;10:124. doi: 10.1186/s13068-017-0796-y. eCollection 2017. Biotechnol Biofuels. 2017. PMID: 28507596 Free PMC article.
-
Draft genome sequence data of Clostridium thermocellum PAL5 possessing high cellulose-degradation ability.Data Brief. 2019 Jul 15;25:104274. doi: 10.1016/j.dib.2019.104274. eCollection 2019 Aug. Data Brief. 2019. PMID: 31406903 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
