

α7nAchR/NMDAR coupling affects NMDAR function and object recognition
- PMID: 24360204
- PMCID: PMC3878138
- DOI: 10.1186/1756-6606-6-58
α7nAchR/NMDAR coupling affects NMDAR function and object recognition
Abstract
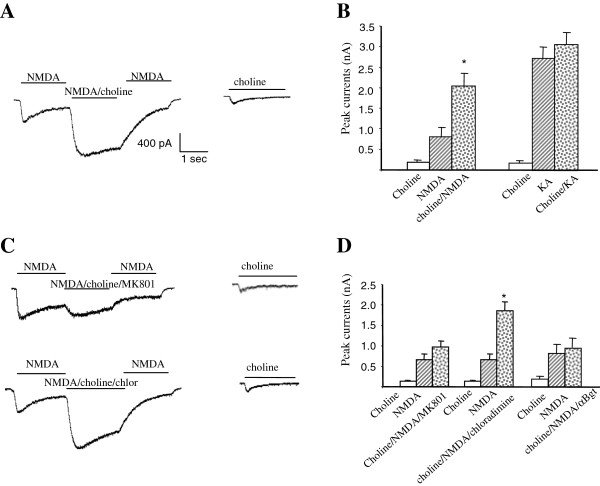
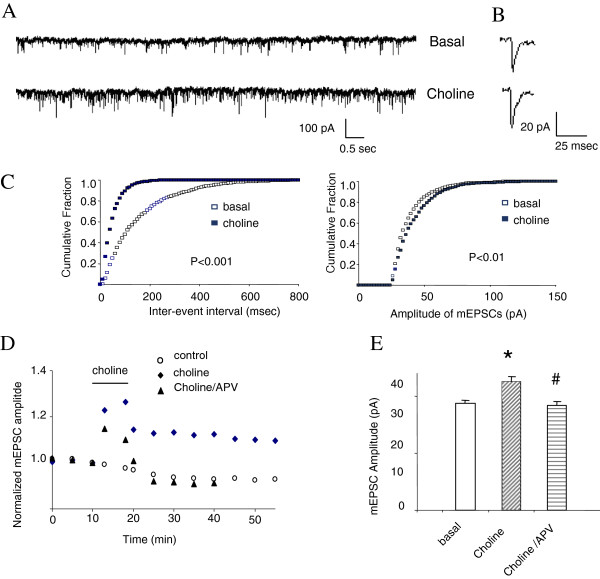
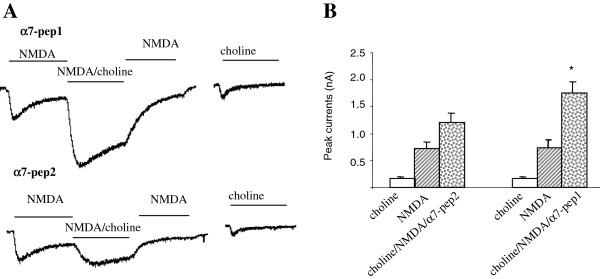
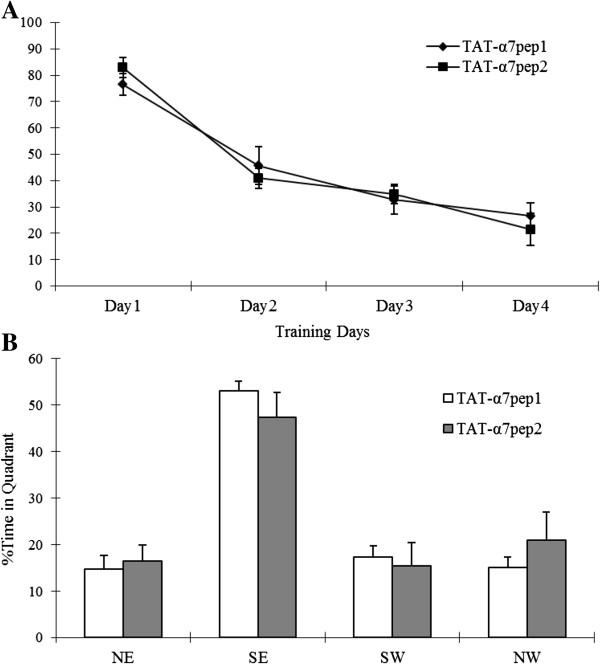
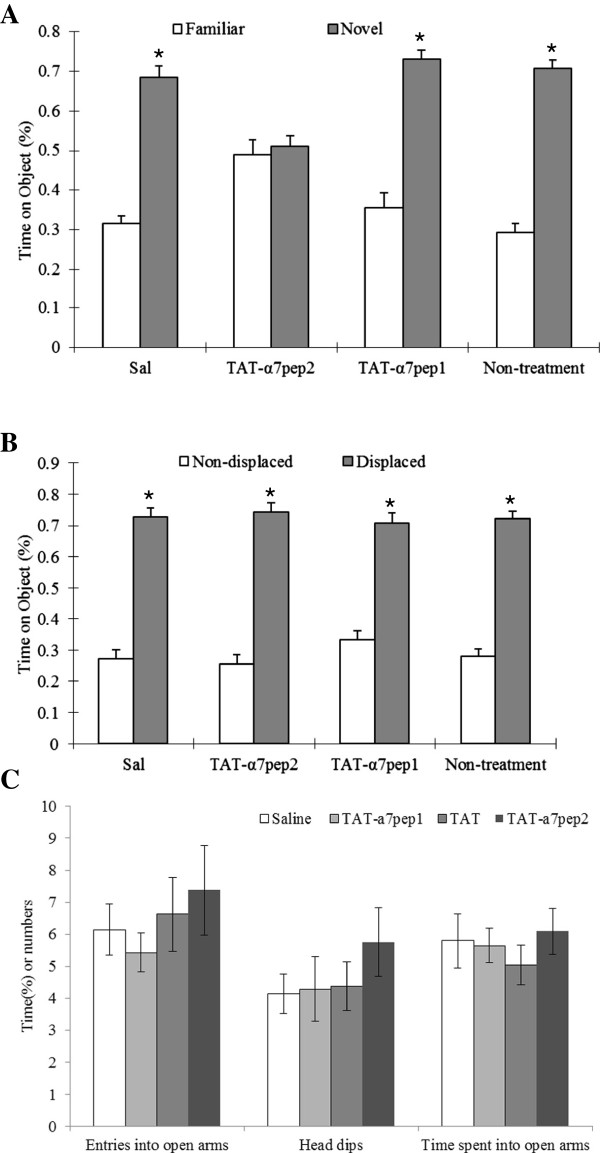
The α7 nicotinic acetylcholine receptor (nAchR) and NMDA glutamate receptor (NMDAR) are both ligand-gated ion channels permeable to Ca2+ and Na+. Previous studies have demonstrated functional modulation of NMDARs by nAchRs, although the molecular mechanism remains largely unknown. We have previously reported that α7nAchR forms a protein complex with the NMDAR through a protein-protein interaction. We also developed an interfering peptide that is able to disrupt the α7nAchR-NMDAR complex and blocks cue-induced reinstatement of nicotine-seeking in rat models of relapse. In the present study, we investigated whether the α7nAchR-NMDAR interaction is responsible for the functional modulation of NMDAR by α7nAchR using both electrophysiological and behavioral tests. We have found that activation of α7nAchR upregulates NMDAR-mediated whole cell currents and LTP of mEPSC in cultured hippocampal neurons, which can be abolished by the interfering peptide that disrupts the α7nAchR-NMDAR interaction. Moreover, administration of the interfering peptide in mice impairs novel object recognition but not Morris water maze performance. Our results suggest that α7nAchR/NMDAR coupling may selectively affect some aspects of learning and memory.
Figures

References
-
- Bartlett TE, Wang YT. The intersections of NMDAR-dependent synaptic plasticity and cell survival. Neuropharmacology. 2013;6:59–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
