

Brd4 and JMJD6-associated anti-pause enhancers in regulation of transcriptional pause release
- PMID: 24360279
- PMCID: PMC3886918
- DOI: 10.1016/j.cell.2013.10.056
Brd4 and JMJD6-associated anti-pause enhancers in regulation of transcriptional pause release
Abstract
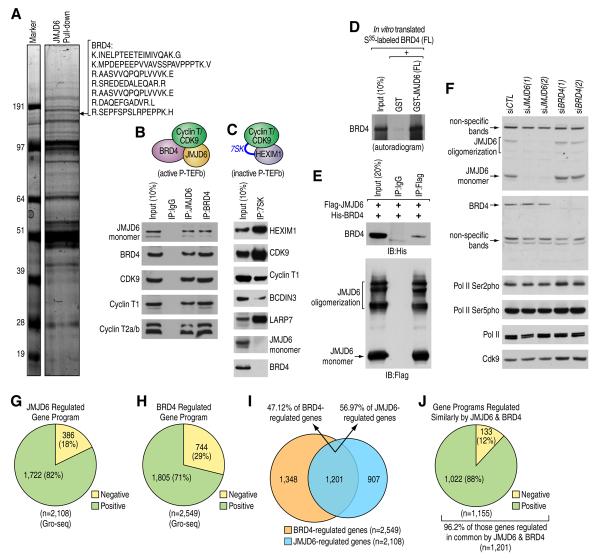
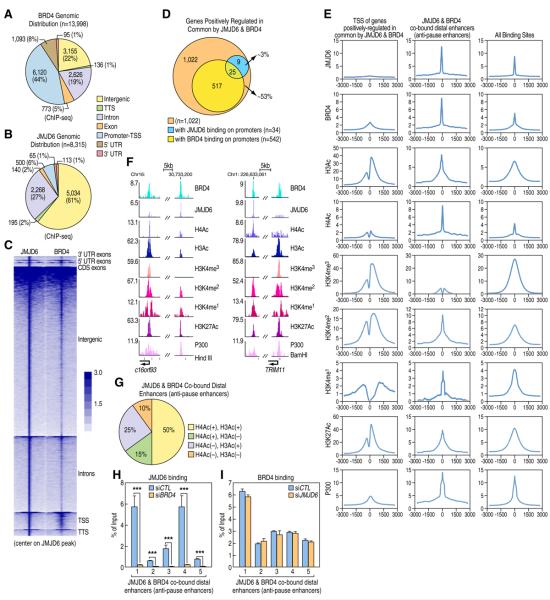
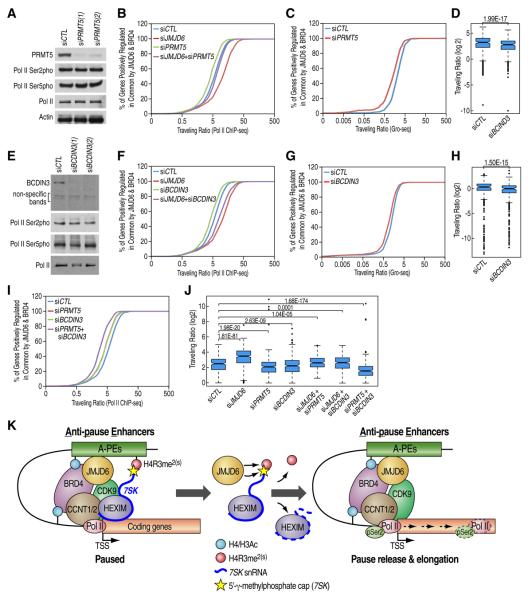
Distal enhancers characterized by the H3K4me(1) mark play critical roles in developmental and transcriptional programs. However, potential roles of specific distal regulatory elements in regulating RNA polymerase II (Pol II) promoter-proximal pause release remain poorly investigated. Here, we report that a unique cohort of jumonji C-domain-containing protein 6 (JMJD6) and bromodomain-containing protein 4 (Brd4) cobound distal enhancers, termed anti-pause enhancers (A-PEs), regulate promoter-proximal pause release of a large subset of transcription units via long-range interactions. Brd4-dependent JMJD6 recruitment on A-PEs mediates erasure of H4R3me(2(s)), which is directly read by 7SK snRNA, and decapping/demethylation of 7SK snRNA, ensuring the dismissal of the 7SK snRNA/HEXIM inhibitory complex. The interactions of both JMJD6 and Brd4 with the P-TEFb complex permit its activation and pause release of regulated coding genes. The functions of JMJD6/ Brd4-associated dual histone and RNA demethylase activity on anti-pause enhancers have intriguing implications for these proteins in development, homeostasis, and disease.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 NS034934/NS/NINDS NIH HHS/United States
- R37 DK039949/DK/NIDDK NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- DK18477/DK/NIDDK NIH HHS/United States
- P41 RR011823/RR/NCRR NIH HHS/United States
- R01 DK018477/DK/NIDDK NIH HHS/United States
- P30 CA023100/CA/NCI NIH HHS/United States
- R01 DK039949/DK/NIDDK NIH HHS/United States
- CA173903/CA/NCI NIH HHS/United States
- DK097748/DK/NIDDK NIH HHS/United States
- NS34934/NS/NINDS NIH HHS/United States
- P01 DK074868/DK/NIDDK NIH HHS/United States
- R01 CA173903/CA/NCI NIH HHS/United States
- HL065445/HL/NHLBI NIH HHS/United States
- R01 HL065445/HL/NHLBI NIH HHS/United States
- DK39949/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
