

The C. elegans CSR-1 argonaute pathway counteracts epigenetic silencing to promote germline gene expression
- PMID: 24360782
- PMCID: PMC3954781
- DOI: 10.1016/j.devcel.2013.11.014
The C. elegans CSR-1 argonaute pathway counteracts epigenetic silencing to promote germline gene expression
Abstract
Organisms can develop adaptive sequence-specific immunity by reexpressing pathogen-specific small RNAs that guide gene silencing. For example, the C. elegans PIWI-Argonaute/piwi-interacting RNA (piRNA) pathway recruits RNA-dependent RNA polymerase (RdRP) to foreign sequences to amplify a transgenerational small-RNA-induced epigenetic silencing signal (termed RNAe). Here, we provide evidence that, in addition to an adaptive memory of silenced sequences, C. elegans can also develop an opposing adaptive memory of expressed/self-mRNAs. We refer to this mechanism, which can prevent or reverse RNAe, as RNA-induced epigenetic gene activation (RNAa). We show that CSR-1, which engages RdRP-amplified small RNAs complementary to germline-expressed mRNAs, is required for RNAa. We show that a transgene with RNAa activity also exhibits accumulation of cognate CSR-1 small RNAs. Our findings suggest that C. elegans adaptively acquires and maintains a transgenerational CSR-1 memory that recognizes and protects self-mRNAs, allowing piRNAs to recognize foreign sequences innately, without the need for prior exposure
Figures
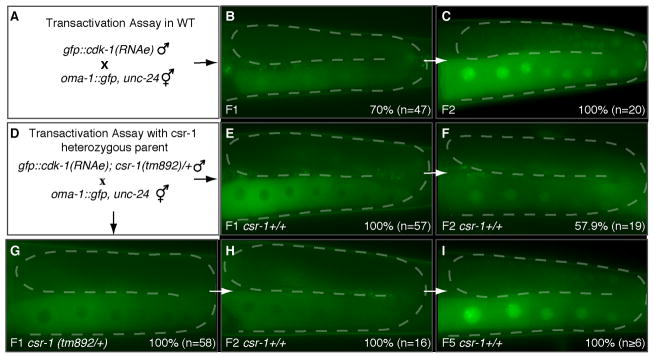
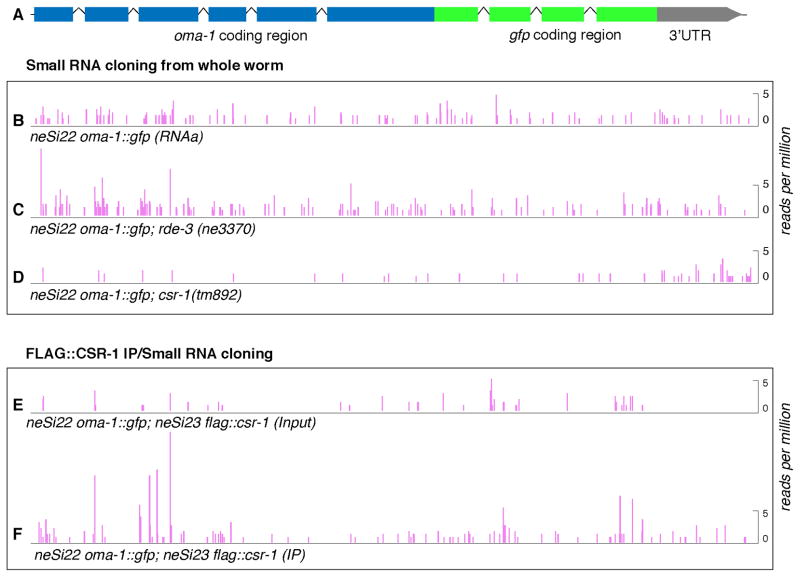
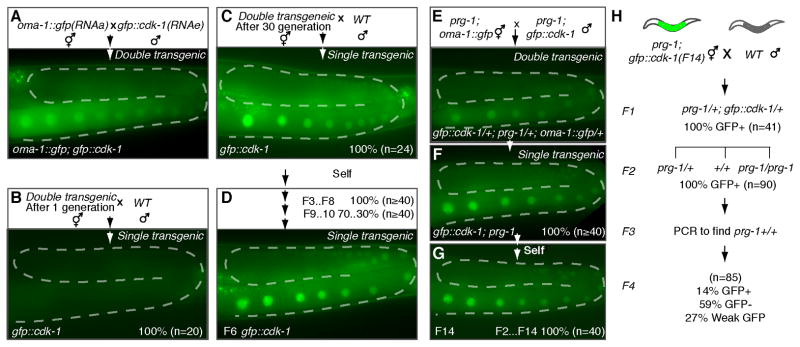
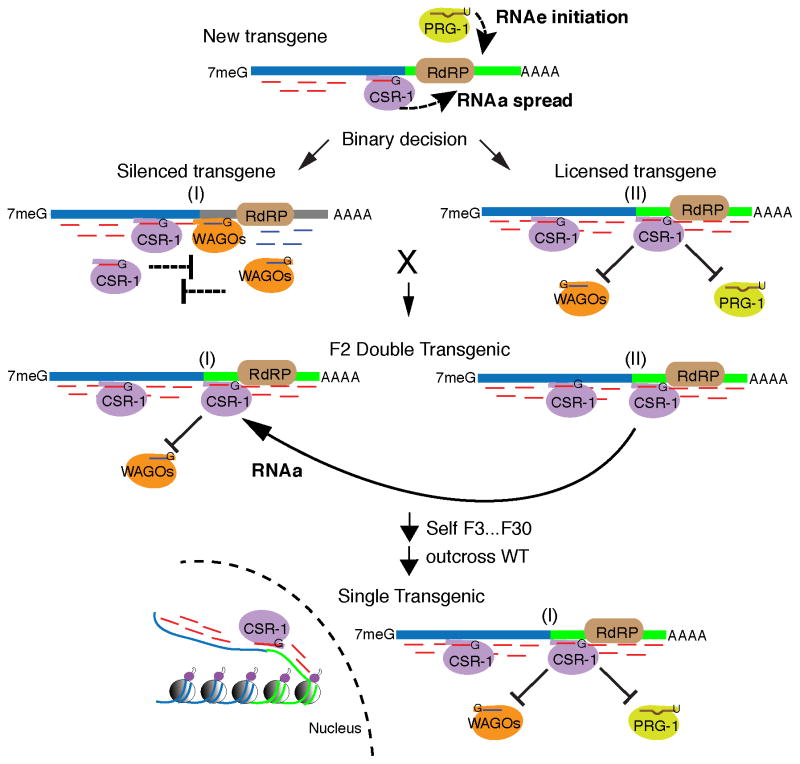
Comment in
-
Is this mine? small RNAs help to decide.Dev Cell. 2013 Dec 23;27(6):599-601. doi: 10.1016/j.devcel.2013.12.005. Dev Cell. 2013. PMID: 24369831
References
-
- Axtell MJ, Jan C, Rajagopalan R, Bartel DP. A two-hit trigger for siRNA biogenesis in plants. Cell. 2006;127:565–577. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
