

A noncoding expansion in EIF4A3 causes Richieri-Costa-Pereira syndrome, a craniofacial disorder associated with limb defects
- PMID: 24360810
- PMCID: PMC3882729
- DOI: 10.1016/j.ajhg.2013.11.020
A noncoding expansion in EIF4A3 causes Richieri-Costa-Pereira syndrome, a craniofacial disorder associated with limb defects
Abstract
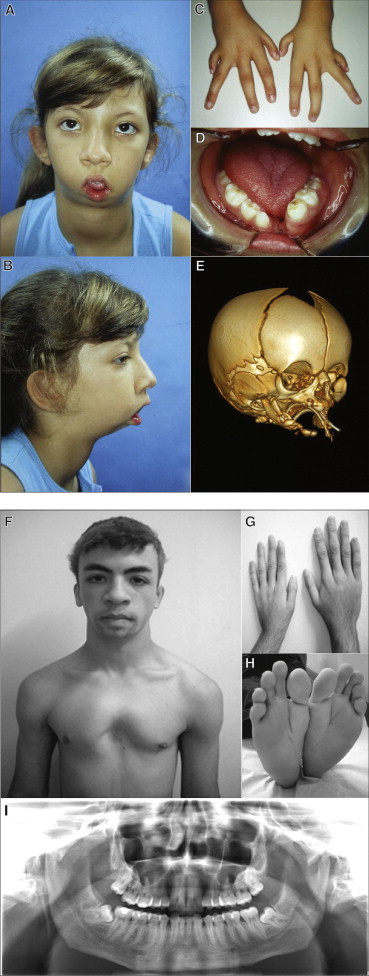
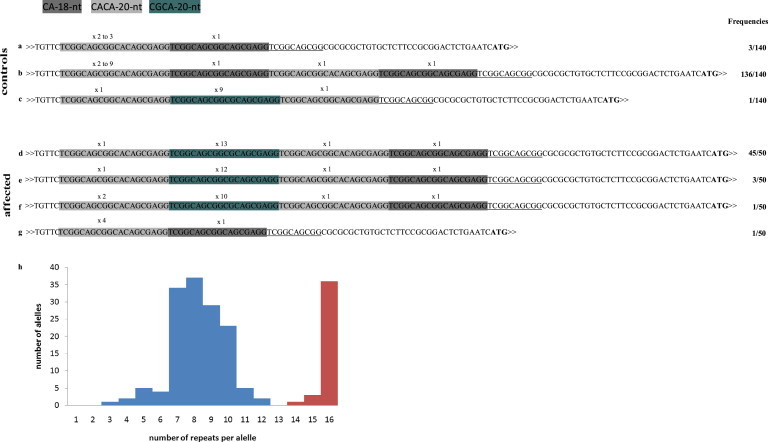
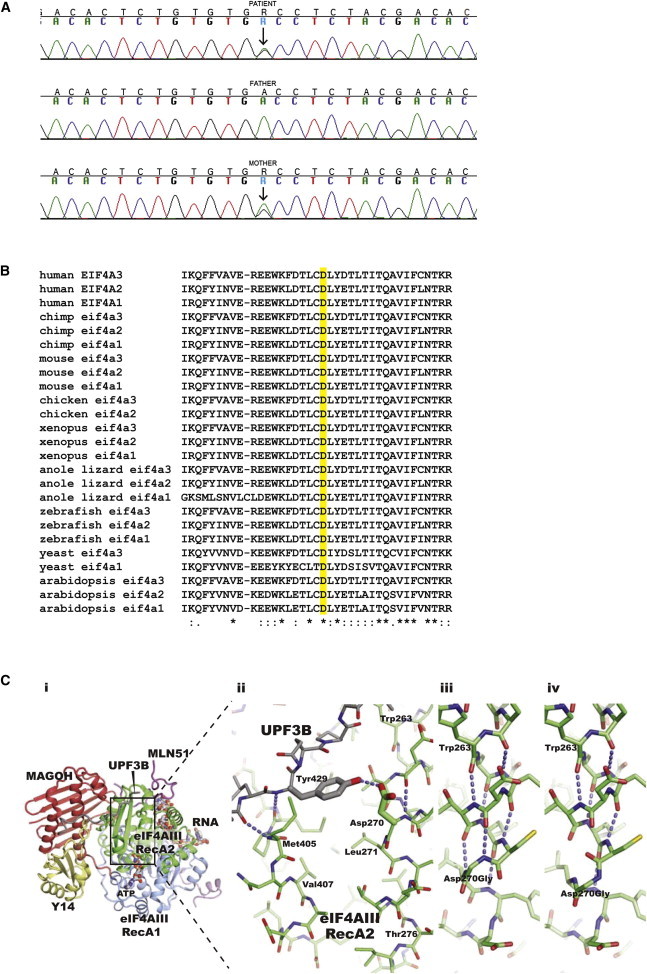
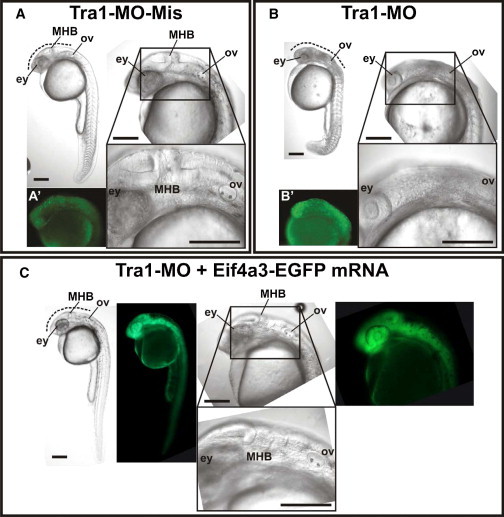
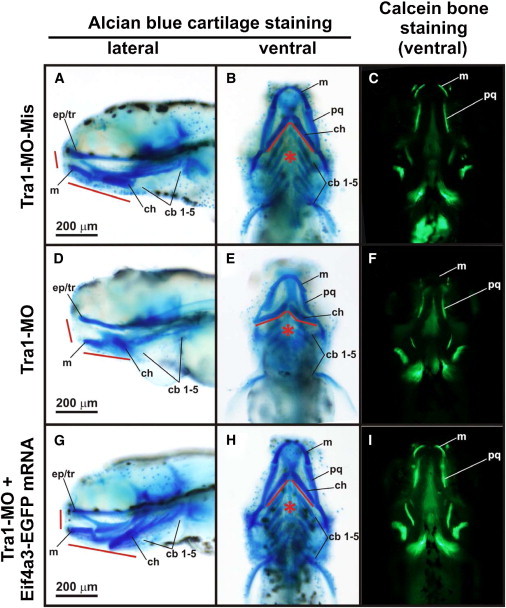
Richieri-Costa-Pereira syndrome is an autosomal-recessive acrofacial dysostosis characterized by mandibular median cleft associated with other craniofacial anomalies and severe limb defects. Learning and language disabilities are also prevalent. We mapped the mutated gene to a 122 kb region at 17q25.3 through identity-by-descent analysis in 17 genealogies. Sequencing strategies identified an expansion of a region with several repeats of 18- or 20-nucleotide motifs in the 5' untranslated region (5' UTR) of EIF4A3, which contained from 14 to 16 repeats in the affected individuals and from 3 to 12 repeats in 520 healthy individuals. A missense substitution of a highly conserved residue likely to affect the interaction of eIF4AIII with the UPF3B subunit of the exon junction complex in trans with an expanded allele was found in an unrelated individual with an atypical presentation, thus expanding mutational mechanisms and phenotypic diversity of RCPS. EIF4A3 transcript abundance was reduced in both white blood cells and mesenchymal cells of RCPS-affected individuals as compared to controls. Notably, targeting the orthologous eif4a3 in zebrafish led to underdevelopment of several craniofacial cartilage and bone structures, in agreement with the craniofacial alterations seen in RCPS. Our data thus suggest that RCPS is caused by mutations in EIF4A3 and show that EIF4A3, a gene involved in RNA metabolism, plays a role in mandible, laryngeal, and limb morphogenesis.
Copyright © 2014 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Favaro F.P., Zechi-Ceide R.M., Alvarez C.W., Maximino L.P., Antunes L.F.B.B., Richieri-Costa A., Guion-Almeida M.L. Richieri-Costa-Pereira syndrome: a unique acrofacial dysostosis type. An overview of the Brazilian cases. Am. J. Med. Genet. A. 2011;155A:322–331. - PubMed
-
- Ferreira de Lima R.L., Moretti-Ferreira D., Richieri-Costa A., Murray J.C. Identity by descent and candidate gene mapping of Richieri-Costa and Pereira syndrome. Am. J. Med. Genet. A. 2003;122A:56–58. - PubMed
-
- Abecasis G.R., Cherny S.S., Cookson W.O., Cardon L.R. Merlin—rapid analysis of dense genetic maps using sparse gene flow trees. Nat. Genet. 2002;30:97–101. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
