

In vivo direct reprogramming of reactive glial cells into functional neurons after brain injury and in an Alzheimer's disease model
- PMID: 24360883
- PMCID: PMC3967760
- DOI: 10.1016/j.stem.2013.12.001
In vivo direct reprogramming of reactive glial cells into functional neurons after brain injury and in an Alzheimer's disease model
Abstract
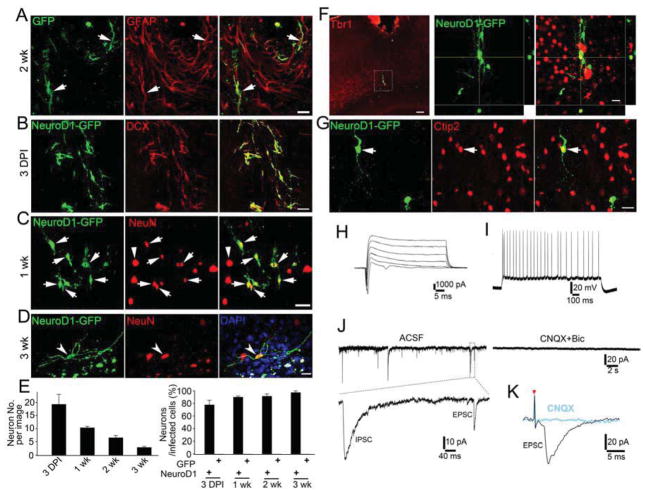
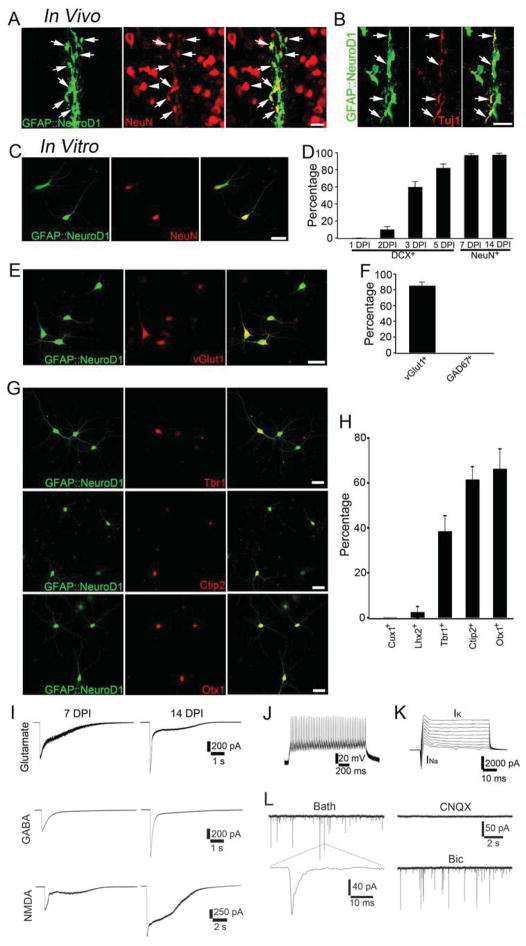
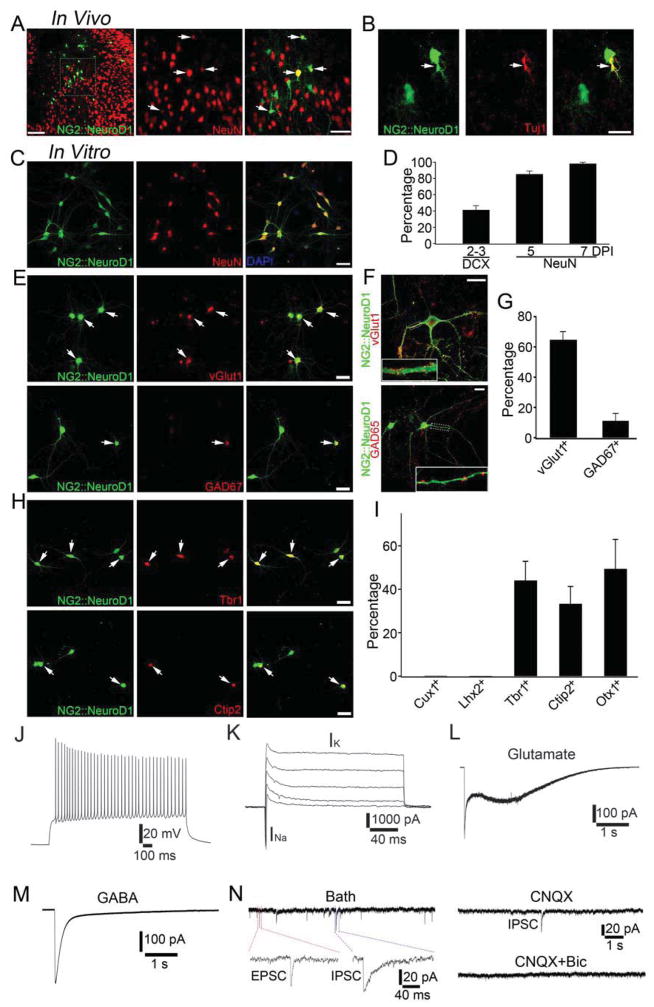
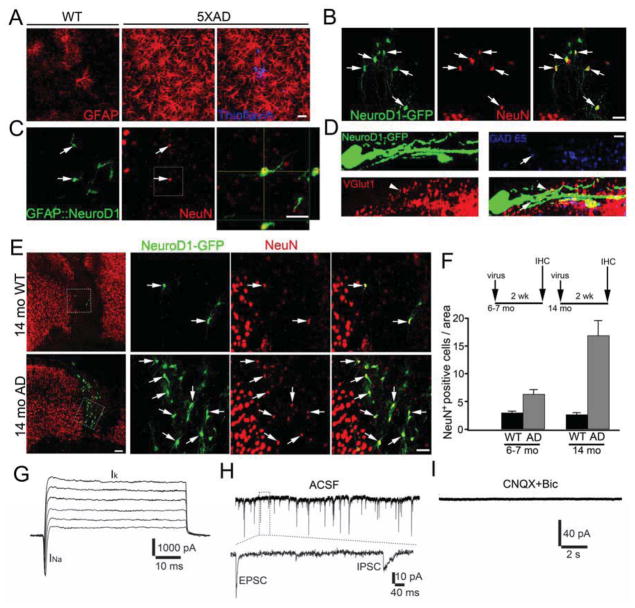
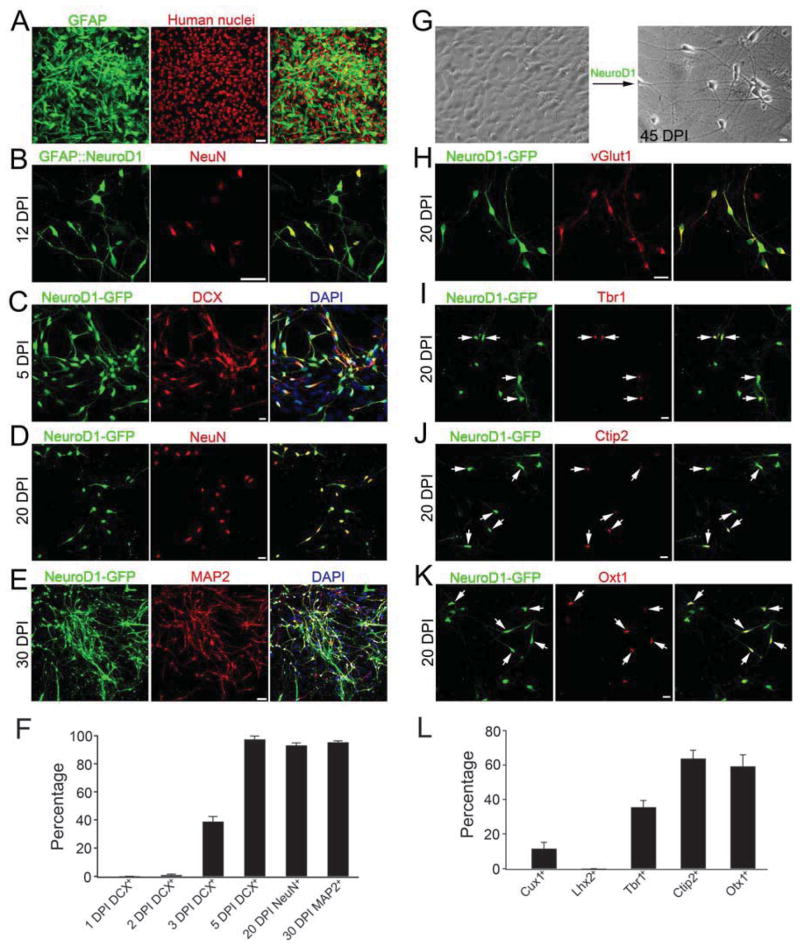
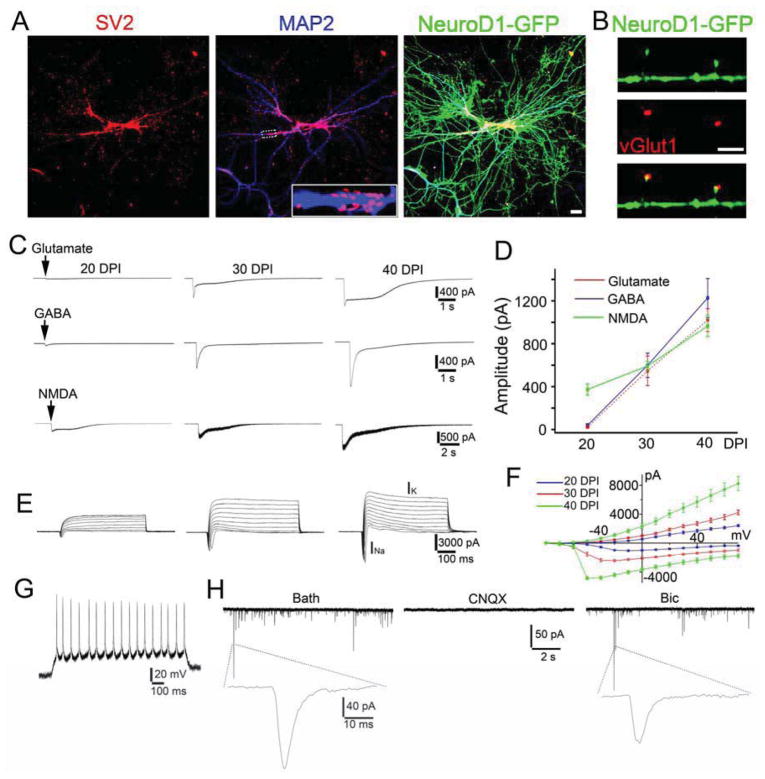
Loss of neurons after brain injury and in neurodegenerative disease is often accompanied by reactive gliosis and scarring, which are difficult to reverse with existing treatment approaches. Here, we show that reactive glial cells in the cortex of stab-injured or Alzheimer's disease (AD) model mice can be directly reprogrammed into functional neurons in vivo using retroviral expression of a single neural transcription factor, NeuroD1. Following expression of NeuroD1, astrocytes were reprogrammed into glutamatergic neurons, while NG2 cells were reprogrammed into glutamatergic and GABAergic neurons. Cortical slice recordings revealed both spontaneous and evoked synaptic responses in NeuroD1-converted neurons, suggesting that they integrated into local neural circuits. NeuroD1 expression was also able to reprogram cultured human cortical astrocytes into functional neurons. Our studies therefore suggest that direct reprogramming of reactive glial cells into functional neurons in vivo could provide an alternative approach for repair of injured or diseased brain.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Turning reactive glia into functional neurons in the brain.Cell Stem Cell. 2014 Feb 6;14(2):133-4. doi: 10.1016/j.stem.2014.01.010. Cell Stem Cell. 2014. PMID: 24506877 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
