

A nuclear perspective on RNAi pathways in metazoans
- PMID: 24361586
- PMCID: PMC3951611
- DOI: 10.1016/j.bbagrm.2013.11.009
A nuclear perspective on RNAi pathways in metazoans
Abstract
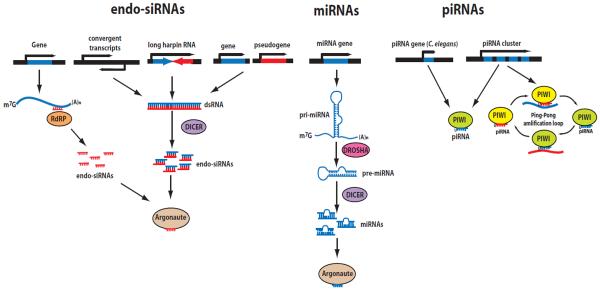
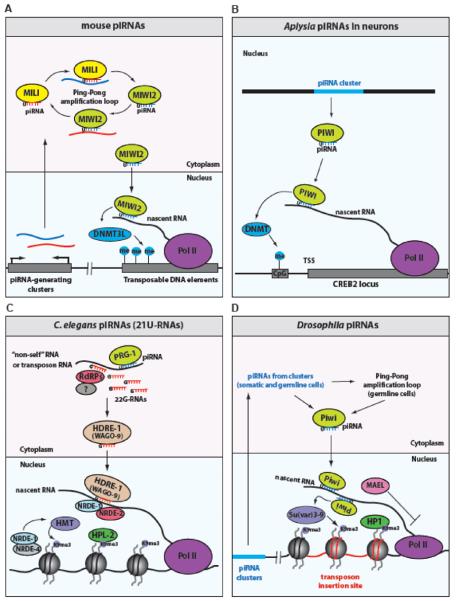
The role of RNA interference (RNAi) in post-transcriptional regulation of complementary targets is well known. However, less is known about transcriptional silencing mechanisms mediated by RNAi. Such mechanisms have been characterized in yeast and plants, which suggests that similar RNA silencing mechanisms might operate in animals. A growing amount of experimental evidence indicates that short RNAs and their co-factor Argonaute proteins can regulate many nuclear processes in metazoans. PIWI-interacting RNAs (piRNAs) initiate transcriptional silencing of transposable elements, which leads to heterochromatin formation and/or DNA methylation. In addition, Argonaute proteins and short RNAs directly regulate Pol II transcription and splicing of euchromatic protein-coding genes and also affect genome architecture. Therefore, RNAi pathways can have a profound global impact on the transcriptional programs in cells during animal development. This article is part of a Special Issue entitled: Chromatin and epigenetic regulation of animal development.
Keywords: Argonaute; Chromatin; RNAi; Transcriptional regulation; endo-siRNAs; piRNAs.
Copyright © 2013 Elsevier B.V. All rights reserved.
Figures
References
-
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. - PubMed
-
- Cogoni C, Romano N, Macino G. Suppression of gene expression by homologous transgenes. Antonie van Leeuwenhoek. 1994;65:205–209. - PubMed
-
- Pal-Bhadra M, Bhadra U, Birchler JA. Cosuppression in Drosophila: gene silencing of Alcohol dehydrogenase by white-Adh transgenes is Polycomb dependent. Cell. 1997;90:479–490. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
