

Adult neural stem cells in distinct microdomains generate previously unknown interneuron types
- PMID: 24362763
- PMCID: PMC4100623
- DOI: 10.1038/nn.3610
Adult neural stem cells in distinct microdomains generate previously unknown interneuron types
Abstract
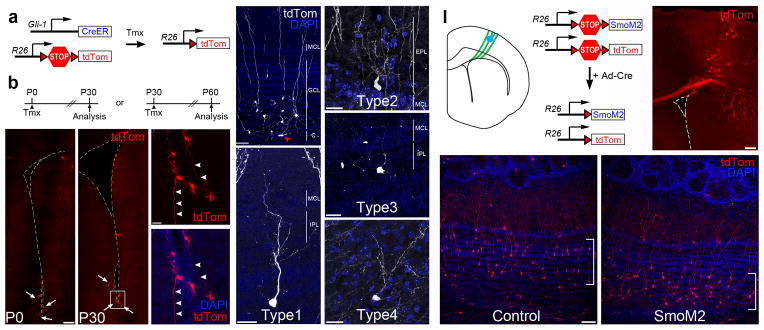
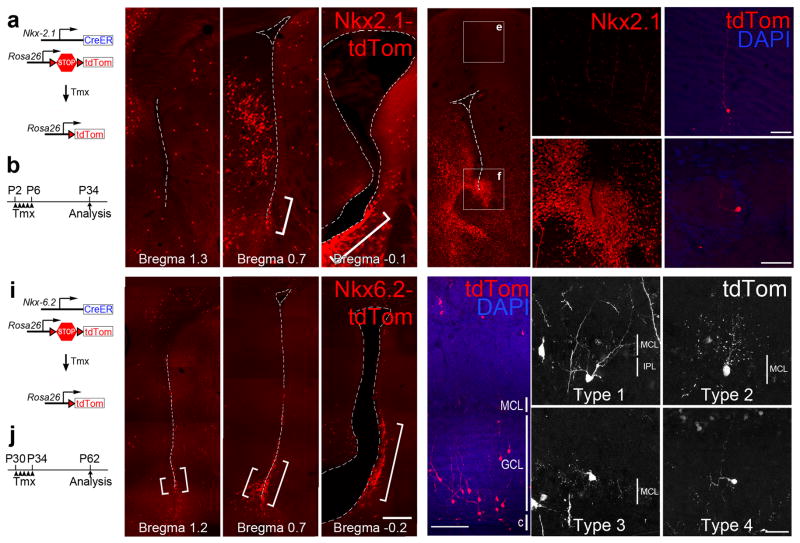
Throughout life, neural stem cells (NSCs) in different domains of the ventricular-subventricular zone (V-SVZ) of the adult rodent brain generate several subtypes of interneurons that regulate the function of the olfactory bulb. The full extent of diversity among adult NSCs and their progeny is not known. Here, we report the generation of at least four previously unknown olfactory bulb interneuron subtypes that are produced in finely patterned progenitor domains in the anterior ventral V-SVZ of both the neonatal and adult mouse brain. Progenitors of these interneurons are responsive to sonic hedgehog and are organized into microdomains that correlate with the expression domains of the Nkx6.2 and Zic family of transcription factors. This work reveals an unexpected degree of complexity in the specification and patterning of NSCs in the postnatal mouse brain.
Figures





References
-
- Imayoshi I, et al. Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nature Neuroscience. 2008;11:1153–1161. - PubMed
-
- Kosaka K, et al. Chemically defined neuron groups and their subpopulations in the glomerular layer of the rat main olfactory bulb. Neurosci Res. 1995;23:73–88. - PubMed
-
- Price JL, Powell TP. The mitral and short axon cells of the olfactory bulb. J Cell Sci. 1970;7:631–651. - PubMed
