

Host-cell sensors for Plasmodium activate innate immunity against liver-stage infection
- PMID: 24362933
- PMCID: PMC4096771
- DOI: 10.1038/nm.3424
Host-cell sensors for Plasmodium activate innate immunity against liver-stage infection
Abstract
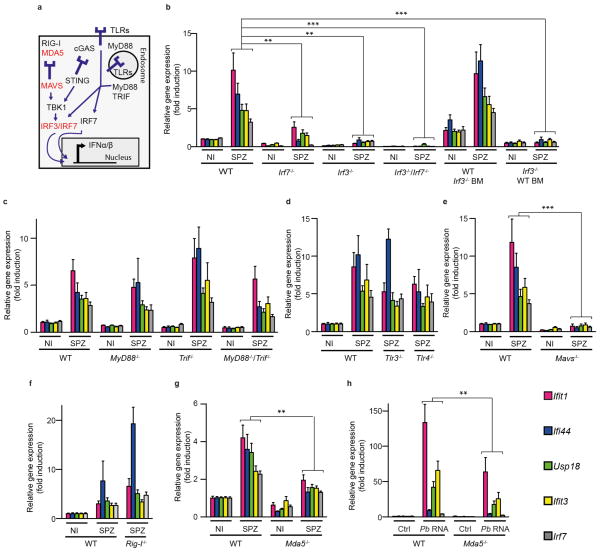
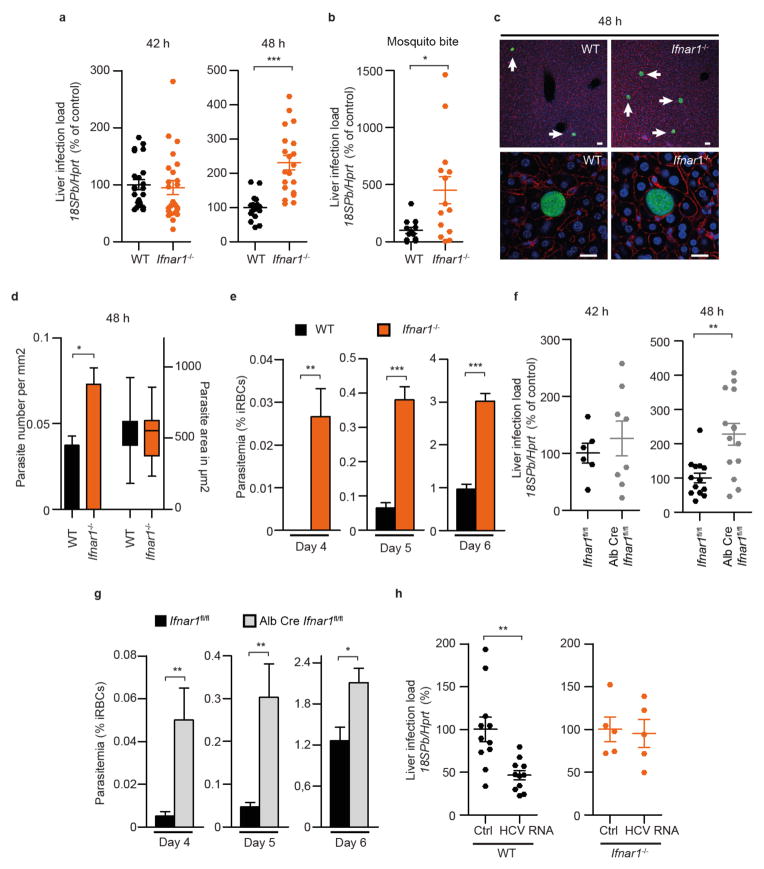
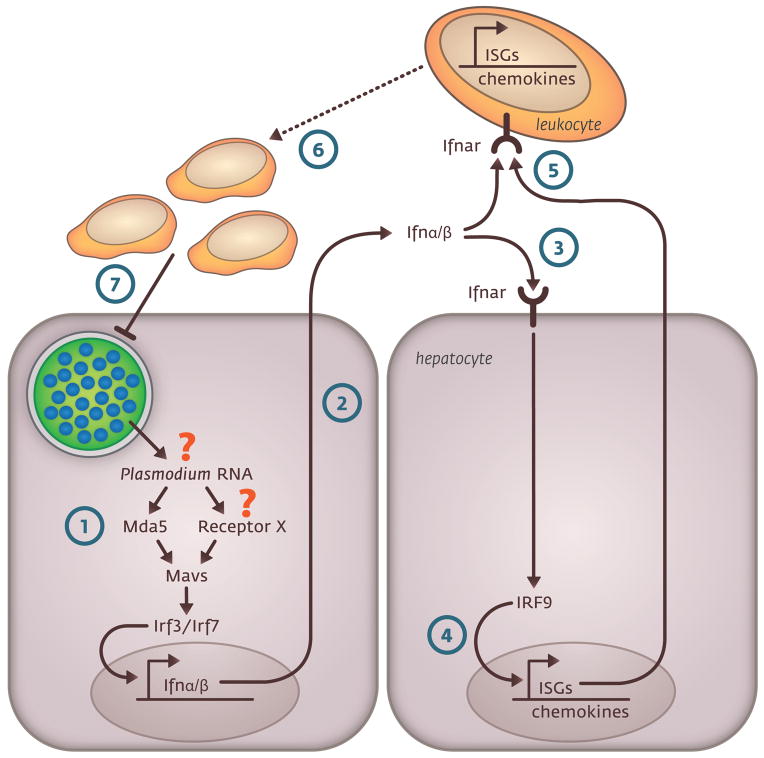
Before they infect red blood cells and cause malaria, Plasmodium parasites undergo an obligate and clinically silent expansion phase in the liver that is supposedly undetected by the host. Here, we demonstrate the engagement of a type I interferon (IFN) response during Plasmodium replication in the liver. We identified Plasmodium RNA as a previously unrecognized pathogen-associated molecular pattern (PAMP) capable of activating a type I IFN response via the cytosolic pattern recognition receptor Mda5. This response, initiated by liver-resident cells through the adaptor molecule for cytosolic RNA sensors, Mavs, and the transcription factors Irf3 and Irf7, is propagated by hepatocytes in an interferon-α/β receptor-dependent manner. This signaling pathway is critical for immune cell-mediated host resistance to liver-stage Plasmodium infection, which we find can be primed with other PAMPs, including hepatitis C virus RNA. Together, our results show that the liver has sensor mechanisms for Plasmodium that mediate a functional antiparasite response driven by type I IFN.
Conflict of interest statement
The authors declare no competing financial interests.
Figures


Comment in
-
Hepatocytes break the silence during liver-stage malaria.Nat Med. 2014 Jan;20(1):17-9. doi: 10.1038/nm.3446. Nat Med. 2014. PMID: 24398958 No abstract available.
References
-
- Prudêncio M, Rodriguez A, Mota MM. The silent path to thousands of merozoites: the Plasmodium liver stage. Nat Rev Microbiol. 2006;4:849–856. - PubMed
-
- Pamplona A, Hanscheid T, Epiphanio S, Mota MM, Vigario AM. Cerebral malaria and the hemolysis/methemoglobin/heme hypothesis: shedding new light on an old disease. Int J Biochem Cell Biol. 2009;41:711–716. - PubMed
-
- Liehl P, Mota MM. Innate recognition of malarial parasites by mammalian hosts. Int J Parasitol. 2012;42:557–566. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
